ช่องІнніและїх budova เมมเบรนชีวภาพและช่องไอออน การเปิดเผยที่สำคัญเกี่ยวกับโครงสร้างและหน้าที่ของช่องไอออน
พลังของช่องไอออนิก
หัวกะทิ - ce vibirkova เพิ่มการเจาะ IC สำหรับการร้องเพลงไอออน สำหรับไอออนอื่นๆ การเจาะจะลดลง ความสั่นสะเทือนดังกล่าวเกิดจากตัวกรองแบบเลือกสรร ซึ่งเป็นส่วนที่สำคัญที่สุดของความพรุนของช่อง ฟิลเตอร์ ครีมของกรีดแคบ อาจจะท้องถิ่น ค่าไฟฟ้า. ตัวอย่างเช่น ช่องสัญญาณที่เลือกได้ของไอออนบวกจะดังขึ้นในบริเวณตัวกรองแบบเลือกเฟ้นที่มีประจุลบของกรดอะมิโนส่วนเกินในคลังของโมเลกุลโปรตีน เพื่อดึงดูดไอออนบวกบวกและปล่อยประจุลบโดยไม่ผ่านรูพรุน
การเจาะ Kerovana - การสร้างІХ vіdkrivatisyaหรือขดตัวเพื่อร้องเพลง keruchih vplivіvในช่อง ช่องปิดอาจลดการเจาะและช่องเปิด - เพิ่มขึ้น เพื่อประโยชน์ด้านพลังงาน IC สามารถจำแนกได้เป็น fallow ได้หลายวิธี เช่น ตัวกระตุ้นศักย์ไฟฟ้า ตัวกระตุ้นด้วยแกนด์ เป็นต้น
การปิดใช้งาน - วัตถุประสงค์ของการสร้าง ІH ในเวลาหนึ่งวันหลังจากการเปิด ลดการแทรกซึมโดยอัตโนมัติจนถึงจุดที่เป็นปัจจัยกระตุ้น ซึ่งกำลังสั่นสะเทือน และทำงานต่อไป การปิดใช้งาน Shvidka เป็นกระบวนการพิเศษที่มีกลไกพิเศษของตัวเองซึ่งดำเนินการในกรณีที่ปิดช่องสัญญาณอย่างเหมาะสม (ปิดใช้งานทั่วไป) การปิด (ปิดใช้งานโดยสมบูรณ์) ของช่องสัญญาณเป็นสิ่งจำเป็นในการประมวลผลกระบวนการ ซึ่งขนานกับกระบวนการ เพื่อให้พวกเขาได้ให้ข้อมูลที่จำเป็น rahunok เปลี่ยนโครงสร้างของช่องโปรตีน และแกนตัวอย่างเช่นที่ช่องที่มีศักยภาพเปิดใช้งานของชาวสวีเดนการปิดใช้งานนั้นใช้สำหรับความช่วยเหลือของ "ซีลก๊อก" โมเลกุลพิเศษซึ่งวางจุกบนมีดหมอราวกับว่าส่งเสียงผู้ชนะที่ห้องอาบน้ำ . ปลั๊กนี้เป็นห่วงกรดอะมิโน (โพลีเปปไทด์) ที่มีเหงื่อออกที่ส่วนท้ายของ มองสามกรดอะมิโนซึ่ง zakaetsya vnutrishne girlo ช่องทางจากด้านข้างของไซโตพลาสซึม Саме тому потенціал-залежні ІЧ для натрію, що забезпечують розвиток потенціалу дії та рух нервового імпульсу, можуть пропускати в клітину іони натрію тільки протягом декількох мілісекунд, а потім вони автоматично закриваються своїми молекулярними пробками, незважаючи на те, що деполяризація, що відкриває, продовжує เด็ก. กลไกอื่นของการปิดใช้งาน IC อาจเป็นการปรับเปลี่ยนช่องสัญญาณคลิตินัสภายในโดยหน่วยย่อยเพิ่มเติม
การปิดกั้นเป็นเป้าหมายของการสร้าง ІCh pіd dієyu ของตัวบล็อกคำพูดเพื่อแก้ไขให้เป็นหนึ่งในค่ายของคุณและไม่ตอบสนองต่อzvichaynіkeruuchіії ในสถานการณ์เช่นนี้ ช่องจะหยุดให้ข้อเสนอแนะเกี่ยวกับข้อความสำคัญ การปิดกั้นการโทรสำหรับตัวบล็อกคำพูด ซึ่งสามารถเรียกได้ว่าเป็นปฏิปักษ์ ตัวบล็อก หรือตัวบล็อก คู่อริ - บรรทัดคำพูดที่เปลี่ยนการเปิดใช้งานสุนทรพจน์อื่นเป็น IC ดังนั้นการกล่าวสุนทรพจน์ของอาคารจึงเชื่อมโยงอย่างดีกับเซลล์รับ ІCh แต่ไม่ใช่อาคารที่เปลี่ยนสถานีเป็นช่องสัญญาณและการตอบสนองของเสียง ออกจากการปิดล้อมของตัวรับทันที iz - การปิดล้อมของ ІCh โปรดทราบว่าการเป็นปรปักษ์กันไม่ obov'yazkovo ทำให้เกิดการปิดล้อมของตัวรับและ yogo ІCh แบบเดียวกัน กลิ่นเหม็นอาจอ่อนลงและ ingibuvati (priignichuvati) น้อยลงในช่องของหุ่นยนต์ แต่อย่าตำหนิมันอีก Agonist-antagonist - ce speech, yakіอาจมีการฉีดกระตุ้นที่อ่อนแอบนตัวรับ แต่ในขณะเดียวกันก็ขัดขวางการพัฒนาของ rhechovina ภายนอกตามธรรมชาติซึ่งจะต้องควบคุม ตัวบล็อก - cervices ที่ข้ามช่องไอออนของหุ่นยนต์เช่นการทำงานร่วมกันของผู้ไกล่เกลี่ยกับตัวรับโมเลกุลไปยังตัวรับใหม่ยังขัดขวางการควบคุมของช่องซึ่งปิดกั้น ตัวอย่างเช่น anticholinergic blockers บล็อก diyu acetylcholine; norepinephrine กับ adrenaline - adrenoblockers; ฮิสตามีน - ตัวบล็อกฮีสตามี ฯลฯ ตัวบล็อกจำนวนมากถูกใช้เพื่อวัตถุประสงค์ในการรักษาเพื่อเตรียมยา เนื้อเพลงเป็นตัวบล็อกคำนี้เก่ากว่าและได้รับชัยชนะในฐานะคำพ้องความหมายสำหรับตัวบล็อก: cholinolytic, adrenolytic ฯลฯ
ความเป็นพลาสติกคือคุณค่าของการสร้างและการเปลี่ยนแปลงอำนาจของมันลักษณะของมัน กลไกการขยายที่ยิ่งใหญ่ที่สุดของกลไกที่ช่วยรับรองความเป็นพลาสติกคือฟอสโฟรีเลชั่นของกรดอะมิโนของโปรตีนช่องจากด้านในของเมมเบรนโดยเอนไซม์โปรตีนไคเนส ส่วนเกินของฟอสฟอริกของ ATP และ GTP จะถูกเพิ่มเข้าไปในโปรตีนของช่องสัญญาณ - และช่องจะเปลี่ยนกำลังของมัน ตัวอย่างเช่น มันแก้ไขตัวเองในสถานะปิด หรือในทางกลับกัน ในสถานะเปิด
Budov และหน้าที่ของช่องไอออนิก พวกเขา Na + , K + , Ca 2+ , Cl - เจาะตรงกลางเซลล์และป้อนชื่อผ่านช่องพิเศษเติมช่องแม่ Rozmir kanaliv dosit maly (เส้นผ่านศูนย์กลาง 05-07 nm) Rosrahunki แสดงให้เห็นว่าพื้นที่ทั้งหมดของคลองครอบครองส่วนที่ไม่มีนัยสำคัญของพื้นผิว เยื่อหุ้มเซลล์.
หน้าที่ของช่องไอออน ในทางที่ต่างออกไป. ขยายให้มากที่สุด - วิธีการแก้ไขแรงดันไฟฟ้าหรือ "แรงดันแคลมป์" (รูปที่ 2.2) สาระสำคัญของวิธีการคือด้วยความช่วยเหลือของระบบอิเล็กทรอนิกส์พิเศษ กระบวนการนี้สามารถเปลี่ยนแปลงและแก้ไขศักยภาพของเมมเบรนให้อยู่ในระดับที่เท่ากัน ด้วยเหตุนี้ ขนาดของอิออนสตรูมาที่ไหลผ่านเมมเบรนจึงถูกควบคุม หากความต่างศักย์กลายเป็น ตามกฎของโอห์ม ค่าของกระแสจะเป็นสัดส่วนกับค่าการนำไฟฟ้าของช่องไอออน ในขั้นตอนเดียวกันของการสลับขั้ว มีช่องทางอื่นๆ เหล่านั้น ซึ่งเข้าสู่คลิตินตามการไล่ระดับไฟฟ้าเคมี นั่นคือ ไอออนิกดีโพลาไรซ์ ซึ่งทำให้เกิดขั้วคลิติน การเปลี่ยนแปลงของ Qia ได้รับการลงทะเบียนเพื่อขอความช่วยเหลือจาก keruyuchy podsiluvach และถูกส่งผ่านเมมเบรน ดีดไฟฟ้าเท่ากับขนาดแต่ขยายเป็นเส้นตรงของเมมเบรน ionic struma ในกรณีนี้ ความต่างศักย์ของเมมเบรนจะไม่เปลี่ยนแปลง สปิลเน vikoristannyaวิธีการตรึงศักยภาพและตัวบล็อกเฉพาะของช่องไอออนได้นำไปสู่การอนุมัติ ประเภทต่างๆช่องไอออนิกที่เยื่อหุ้มเซลล์
ขณะนี้มีการติดตั้งช่องสัญญาณหลายประเภทสำหรับไอออนต่างๆ (ตารางที่ 2.1) บางตัวมีความเฉพาะเจาะจงมากขึ้น บางตัวสามารถข้ามไอออนอื่นๆ นอกเหนือจากไอออนหลักได้
การสร้างฟังก์ชั่นของช่องสัญญาณให้ได้มากที่สุดโดยวิธีการตรึงเฉพาะที่เพื่อศักยภาพของเส้นทาง ข้าว. 2.3 ก) บีบไมโครอิเล็กโทรด (ไมโครปิเปต) ด้วยสเปรย์เกลือ กดไปที่พื้นผิวของเมมเบรนแล้วสร้างสเปรย์ขนาดเล็ก ในเวลาเดียวกัน ส่วนของเมมเบรนจะเคลื่อนขึ้นไปที่ไมโครอิเล็กโทรด ทันทีที่ตรวจพบช่องไอออนในโซนปิ๊กอัพ กิจกรรมของช่องสัญญาณเดียวจะถูกลงทะเบียน ระบบการล้อเล่นและการลงทะเบียนกิจกรรมไปยังช่องสัญญาณได้รับผลกระทบเพียงเล็กน้อยจากระบบการแก้ไขแรงดันไฟฟ้า
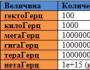
ตาราง 2.1.ช่องไอออนที่สำคัญที่สุดและกระแสไอออนของผู้ตื่นตัว
บันทึก. TEA, เตตระเอทิลแอมโมเนียม; TTX - เตโตรโดท็อกซิน
ส่วนด้านนอกของคลองสามารถเข้าถึงได้บางส่วนในขณะที่ส่วนในของคลองมีความสำคัญ P. G. Kostyuk ได้พัฒนาวิธีการฟอกไตภายในเซลล์ซึ่งช่วยให้การทำงานของโครงสร้างทางเข้าและทางออกของช่องไอออนิกโดยไม่ปิดกั้นไมโครอิเล็กโทรด ปรากฏว่าส่วนหนึ่งของช่องไอออนซึ่งเปิดออกที่ด้านหลังของช่องคอ สำหรับความสามารถในการทำงานของมัน ทำให้แห้งในส่วนของช่องทาง ซึ่งถูกทำให้เป็นสัตว์อยู่ตรงกลางด้านในสุด
ช่องไอออนให้พลังงานเมมเบรนที่สำคัญสองอย่าง ได้แก่ ความสามารถในการคัดเลือกและการนำไฟฟ้า
หัวกะทิ,หรือ ความสั่นสะเทือนช่องสัญญาณถูกยึดด้วยโครงสร้างโปรตีนพิเศษ ช่องสัญญาณจำนวนมากถูกทำให้เป็นไฟฟ้า เพื่อให้อาคารเหล่านี้สามารถฝากไว้ได้ขึ้นอยู่กับขนาดของศักยภาพของเมมเบรน ช่องสัญญาณมีลักษณะการทำงานต่างกัน โดยเฉพาะอย่างยิ่งความสำคัญของโครงสร้างโปรตีน ซึ่งเรียกว่าทางเข้าสู่ช่องและทางออก (ที่เรียกว่า comirn_mechanisms)
5. ความเข้าใจเกี่ยวกับการตื่นตัว พารามิเตอร์ของความตื่นตัวของระบบประสาทและลิ้น: เกณฑ์การหยอกล้อ (rheobase), ชั่วโมงเช้า (chronaxia) การสูญเสียความแข็งแกร่งของ rozdratuvannya ในช่วงเวลาของวัน (เส้นโค้ง Gorweg-Weiss) วัสดุทนไฟ
ความตื่นตัว- Zdatnіst kіtinyvіdpovіdatiบน razdratuvannya molding PD และปฏิกิริยาเฉพาะ
1) เฟสของการสลับขั้วในท้องถิ่น - การสลับขั้วบ่อยครั้งของเมมเบรน (การซึมผ่านของ Na + ใน clitin) ถ้าทาโต๊ะเครื่องแป้งตัวเล็กก็จะแข็งแรง
การสลับขั้วในพื้นที่ - ระยะความสูงส่ง
2) ระยะของการหักเหของแสงสัมบูรณ์ - พลังของเนื้อเยื่อที่ตื่นตัวไม่สร้าง PD สำหรับแรงใดๆ
3) ระยะของการหักเหของแสง
4) ขั้นตอนของการเกิดซ้ำเต็มรูปแบบ - razdratuvannya - การทำซ้ำที่แข็งแกร่งอีกครั้ง
5) เฟสของไฮเปอร์โพลาไรเซชัน - ความตื่นตัวน้อยกว่า (ต่ำกว่าปกติ) สิ่งเร้าอาจมากกว่า
ความสามารถในการทำงาน- การประเมินความตื่นตัวของเนื้อเยื่อผ่านจำนวน PD สูงสุดที่เป็นไปได้ในหนึ่งชั่วโมง
หยุดความเสียหาย:
1) กฎแห่งความแข็งแกร่ง - ความแรงของโพดราซนิกสามารถเป็นธรณีประตูหรือเหนือเกณฑ์ (ค่าต่ำสุดของแรงที่เรียกร้องให้ตื่นตัว) การหยอกล้อที่แข็งแกร่งที่สุดคืออะไรการตื่นที่แข็งแกร่งที่สุด - การรวมตัวของเนื้อเยื่อน้อยลง (stovbur ประสาท, m'yaz, vinyatok - MMC)
2) กฎแห่งชั่วโมง - โพดราซนิกที่ทำให้ร่างกายทรุดโทรมเล็กน้อยอาจเพียงพอสำหรับการปลุกที่มีความผิด
Mіzhโดยแรงของชั่วโมงที่แล้ว, ความรกร้างตามสัดส่วนที่ชายแดนของmіzhตามชั่วโมงขั้นต่ำและโดยแรงขั้นต่ำ ความแรงขั้นต่ำ - rheobase - คือความแข็งแกร่งที่เรียกร้องให้ตื่นขึ้นและนอนในสายลม ชั่วโมงขั้นต่ำคือชั่วโมงสีน้ำเงิน Chronaxia - ความตื่นตัวของเนื้อเยื่ออื่น ๆ เหล่านั้นเป็นเวลาหนึ่งชั่วโมงซึ่งเป็นสาเหตุของการตื่นขึ้น
ยิ่งมีพละกำลังมากเท่าไร ก็ยิ่งมีพลังในการร้องเพลงมากขึ้นเท่านั้น
เจ้าหน้าที่ที่สร้าง WFP:
1) ความแตกต่างของความเข้มข้นของโซเดียมและโพแทสเซียม
2) อัตราการซึมผ่านของโซเดียมและโพแทสเซียม
3) การทำงานของปั๊ม Na-K (3 Na+ ถูกสูบออก, 2 K+ ถูกหมุน)
ความรกร้างระหว่างความแรงของโพดราซนิกและค่าเล็กน้อยของมัน ซึ่งจำเป็นสำหรับปฏิกิริยาขั้นต่ำของปฏิกิริยาของโครงสร้างที่มีชีวิต ดีกว่าที่จะเดินบนเส้นโค้งแรงที่เรียกว่า - ชั่วโมง (เส้นโค้ง Goorweg-Weiss-Lapik)
จากการวิเคราะห์เส้นโค้ง เห็นได้ชัดว่าถึงแม้ความแข็งแกร่งของโพดราซนิกจะแข็งแกร่งมาก ด้วยความไม่สำคัญที่ไม่เพียงพอ จะไม่มีปฏิกิริยาใด ๆ ในการให้ยา (จุดของ levoruch ในคอหนืดของอติพจน์) ปรากฏการณ์ที่คล้ายกันนี้ได้รับการปกป้องสำหรับ trivalo di subthreshold podrazniki ความแรงขั้นต่ำของการดีด (หรือความตึงเครียด) zdatna viklikati zbudzhennya ถูกตั้งชื่อโดย Lapik rheobase (vіdrіzok ordinate OA) ช่วงเวลาที่เล็กที่สุดของชั่วโมง, การยืดของดีดบางชนิด, เท่ากับความแข็งแรงของ undertow reobasy, เรียกออกมาในเนื้อผ้าของการตื่น, เรียกว่า chronaxia (ในกรณีของ abscissa OF) เป็นตัวบ่งชี้ของ ขีด จำกัด ของการล้อเล่น Chronaxia ลดลง δ (หนึ่งในพันของวินาที) ขนาดของโครนาเซียสามารถใช้เพื่อตัดสินความรุนแรงของการกระตุ้นในเนื้อผ้า: ยิ่ง chronaxia น้อยลง ความเร้าอารมณ์ก็จะยิ่งมากขึ้น ความสอดคล้องของเส้นประสาทและเส้นใยเมือกของมนุษย์นั้นเก่าแก่ถึงหนึ่งพันและหนึ่งในพันของวินาที และลำดับของเนื้อเยื่อหลัก เช่น เส้นใยเมือกของถุงคางคก เป็นหนึ่งร้อยของวินาที
ลำดับเหตุการณ์ของเนื้อเยื่อที่ตื่นอยู่นั้นมีความหลากหลายในการทดลองและในสรีรวิทยาของการกีฬาในคลินิก ด้วยความช่วยเหลือของเส้นทางของการบรรเทา chronaxia m'yaza นักประสาทวิทยาสามารถสร้างการปรากฏตัวของการแตกของเส้นประสาท ruch จำเป็นต้องระบุว่าผู้ที่ตกอับสามารถเข้มแข็งได้ แม่เป็นคนไม่สำคัญ แต่ความเร็วต่ำจะเพิ่มขึ้นในชั่วโมงจนถึงค่าเส้นเขตแดน คุณไม่โทษว่าเป็นเพราะโอกาสนั้น สิ่งที่แนบมาของเนื้อเยื่อที่ตื่นขึ้นอย่างเต็มที่ทำให้ชื่อที่พักหายไป ที่พักได้รับผลกระทบจากความจริงที่ว่าในหนึ่งชั่วโมงการเติบโตของความแข็งแรงของชุดชั้นในในเนื้อผ้าจะเพิ่มขึ้น การเปลี่ยนแปลงที่ใช้งานอยู่ซึ่งยกระดับการหยอกล้อและสับเปลี่ยนการพัฒนาของการตื่นขึ้น ในพิธีกรรมดังกล่าว ความเร็วของการล้อเล่นที่เพิ่มขึ้นในชั่วโมงหรือการไล่ระดับของการหยอกล้อมีความสำคัญต่อการปลุกให้ตื่นขึ้น
กฎการพัฒนาเกรเดียนต์ ปฏิกิริยาของการส่องสว่างแบบสดต่อทีเซอร์คือการตกอยู่ในรูปแบบของการไล่ระดับการล้อเล่น ซึ่งเกิดจากคำศัพท์หรือความเยือกเย็นของการเติบโตของทีเซอร์ในหนึ่งชั่วโมง: อะไรคือการไล่ระดับที่ยิ่งใหญ่ที่สุดของการล้อเล่น นี่คือ ปฏิกิริยาที่แข็งแกร่งที่สุด (ในการร้องเพลงอินเตอร์) ในอากาศของการส่องสว่างที่ตื่นขึ้น
จากกฎเดียวกัน การล้อเล่นควรพับระหว่างทีเซอร์และโครงสร้างที่ตื่นตัวสำหรับ її vzaєmodії สำหรับ viniknennya zbudzhennya podraznik มีความผิดในเรื่องความแข็งแกร่งของชายแดนแม่ความเหลื่อมล้ำของแม่และการเติบโตอย่างรวดเร็วของแม่ในชั่วโมงนั้น
6. ปั๊มอิออน (ATP-azy): K+-Na+-eva, Ca2+-eva (พลาสโมเลไมต์ของ sarcoplasmic reticulum), ตัวแลกเปลี่ยน H+–K+
จากผลการวิจัยล่าสุดพบว่าในเยื่อหุ้มชีวภาพมีปั๊มไอออนซึ่งทำงานเพื่อพลังงานอิสระของการไฮโดรไลซิสของ ATP ซึ่งเป็นระบบพิเศษของโปรตีนหนึ่ง (ขนส่ง ATPase)
ปัจจุบันมีปั๊มอิเล็กโทรเจนิกไอออนิกสามประเภทที่ถ่ายเทไอออนผ่านเมมเบรนอย่างแข็งขัน (รูปที่ 13)
การถ่ายโอนไอออนโดยการขนส่ง ATPases เกิดจากกระบวนการถ่ายโอนที่ตามมาด้วยปฏิกิริยาเคมีสำหรับการเผาผลาญพลังงานของคลิติน
ระหว่างการทำงานของ K+-Na+-ATPase สำหรับการไหลของพลังงานซึ่งเปลี่ยนแปลงระหว่างการไฮโดรไลซิสของโมเลกุล ATP ของผิวหนัง โพแทสเซียมไอออนสองตัวจะถูกถ่ายโอนไปยังคลิตินและโซเดียมไอออนสามตัวจะถูกสูบออกจากคลิตินในเวลาเดียวกัน ด้วยวิธีนี้ ความเข้มข้นของโพแทสเซียมไอออนในคลิทินัมจึงลดลง ซึ่งมีความสำคัญทางสรีรวิทยาอย่างมาก
สัญญาณของ "ไบโอปั๊ม":
1. รีบเร่งกับการไล่ระดับศักย์ไฟฟ้าเคมี
2. การไหลของคำพูดของผ้าพันแผลจากการไฮโดรไลซิสของ ATP (หรือแหล่งพลังงานอื่น ๆ )
3. ความไม่สมมาตรของเครื่องขนส่ง
4. การไฮโดรไลซิสของปั๊มในหลอดทดลองในอาคารของ ATP นั้นน้อยกว่าเมื่อมีไอออนเงียบซึ่งสามารถถ่ายโอนในร่างกายได้
5. เมื่อปั๊มถูกติดตั้งเป็นชิ้น ๆ ตรงกลางของเส้นเลือดสามารถประหยัดหัวกะทิ
กลไกระดับโมเลกุลของ ATPase ของหุ่นยนต์ยังไม่ได้รับการแก้ไข ขั้นตอนหลักของกระบวนการพับเอนไซม์นั้นถูกรบกวน ใน K + -Na + -ATPhases ที่แตกต่างกัน มีขั้นตอนของการถ่ายโอนไอออนเนื่องจากการไฮโดรไลซิสของ ATP
แผนภาพแสดงให้เห็นว่าขั้นตอนสำคัญในการทำงานของเอนไซม์คือ:
1) การตรึงเอ็นไซม์คอมเพล็กซ์ด้วย ATP บนพื้นผิวด้านในของเมมเบรน (ปฏิกิริยาถูกกระตุ้นโดยแมกนีเซียมไอออน)
2) ผูกมัดกับไตรออกซิโอนิกโซเดียมคอมเพล็กซ์;
3) ฟอสโฟรีเลชั่นของเอนไซม์สำหรับการแปลงอะดีโนซีนไดฟอสเฟต
4) พลิก (flip-flop) ไปที่เอนไซม์ที่อยู่ตรงกลางของเมมเบรน
5) ปฏิกิริยาของการแลกเปลี่ยนโซเดียมไอออนกับโพแทสเซียมซึ่งทำปฏิกิริยากับผิวด้านนอกของเมมเบรน
6) การพลิกกลับของคอมเพล็กซ์ของเอนไซม์ด้วยการถ่ายโอนโพแทสเซียมไอออนเข้าสู่กลางเซลล์
7) เปลี่ยนเอ็นไซม์ให้กลายเป็น ค่ายพักร้อนถึงระดับโพแทสเซียมไอออนและอนินทรีย์ฟอสเฟต (P)
ด้วยวิธีนี้ สำหรับวัฏจักรสุดท้าย โซเดียมสามไอออนจะถูกปลดปล่อยออกจากเซลล์ โพแทสเซียมไอออนสองตัวจะอุดมไปด้วยไซโทพลาซึม และโมเลกุล ATP หนึ่งโมเลกุลจะถูกไฮโดรไลซ์
ช่องเหล่านี้สร้างขึ้นโดยเฉพาะในเยื่อหุ้มเซลล์ซึ่งเป็นโปรตีนโอลิโกเมอร์ (ซึ่งประกอบด้วยหน่วยย่อยรูปลอก) โมเลกุลโปรตีนเป็นโมเลกุลโปรตีนไปยังช่องกลางเนื่องจากแทรกซึมเมมเบรนในลักษณะที่รูพรุนของช่องก่อตัวขึ้นในศูนย์กลางที่ชอบน้ำผ่านร่องเพื่อเจาะเข้าไปในคลิตินาของอาคารซึ่งมีเส้นผ่านศูนย์กลาง ไม่เกินเส้นผ่านศูนย์กลางของรูขุมขน (เสียง tse ioni)
ใกล้กับส่วนหัวของยูนิตย่อยของช่องสัญญาณ ได้มีการพัฒนาระบบของหน่วยย่อยรูปลอก ดังนั้นจึงสร้างเพลตสำหรับการมีปฏิสัมพันธ์กับโปรตีนควบคุมเมมเบรน ผู้ไกล่เกลี่ยต่างๆ รวมถึงคำปราศรัยที่ออกฤทธิ์ทางเภสัชวิทยา
การจำแนกประเภทของช่องไอออนสำหรับหน้าที่:
1) สำหรับจำนวนไอออนซึ่งช่องสามารถซึมผ่านได้ช่องจะแบ่งออกเป็นส่วนที่เลือก (เจาะเฉพาะไอออนประเภทเดียวเท่านั้น) และแบบไม่คัดเลือก (เจาะทะลุสำหรับชนิดของไอออนรูปลอก)
2) ตามลักษณะของไอออน, yakі กลิ่นเหม็นที่จะส่งต่อ Na +, Ca ++, Cl-, K +-channels;
3) ตามวิธีการควบคุม แบ่งออกเป็นเงินฝากที่มีศักยภาพและอิสระที่อาจเกิดขึ้น ช่องสัญญาณที่ขึ้นกับศักยภาพจะตอบสนองต่อการเปลี่ยนแปลงในศักยภาพของเยื่อคลิติน และเกินศักยภาพที่เข้าถึงได้ของค่าเพลง ช่องจะเปลี่ยนเป็นสถานีที่ทำงานอยู่ โดยค่อยๆ ส่งไอออนจากการไล่ระดับความเข้มข้นของไอออน ดังนั้นช่องโซเดียมและแคลเซียมจึงมีศักยภาพสะสม การกระตุ้นจะเกิดขึ้นเมื่อศักยภาพของเมมเบรนลดลงเหลือ -50-60 mV เมื่อการไหลของไอออน Na + และ Ca ++ ในเลือดลดศักยภาพของยีนสงบ ช่องทางที่สะสมโพแทสเซียมจะถูกกระตุ้นในระหว่างการพัฒนาของ PD I ป้องกันการดีดของไอออน K+ จากเซลล์ และทำให้เกิดการรีโพลาไรเซชันของเมมเบรน
ช่องทางอิสระที่อาจเกิดขึ้นตอบสนองการเปลี่ยนแปลงศักย์ของเมมเบรนต่างกัน แต่กับปฏิกิริยาของตัวรับ เนื่องจากกลิ่นของปฏิกิริยาระหว่างกัน และของลิแกนด์ ดังนั้น Cl-channels จะจับกับตัวรับกรด g-aminobutyric และเมื่อตัวรับกลิ่นเหม็นมีปฏิสัมพันธ์กับมัน พวกมันจะถูกกระตุ้นและให้กระแสของคลอรีนไอออนเข้าสู่ clitina ทำให้เกิดไฮเปอร์โพลาไรเซชันและความตื่นตัวลดลง
3. เมมเบรนที่มีศักยภาพเพื่อความอุ่นใจ
ภาคเรียน "ศักยภาพของเมมเบรนความสงบจิตสงบใจ » เป็นเรื่องปกติที่จะเรียกความแตกต่างของศักยภาพของเมมเบรน ซึ่งอยู่ระหว่างไซโตพลาสซึมและความแตกต่างของเซลล์ที่พบบ่อยที่สุด หากคลิติน (ไฟเบอร์) พักอยู่ที่สถานีของความสงบทางสรีรวิทยา ประจุภายในจะเป็นลบเมื่อเทียบกับเสียง ซึ่งเป็นที่ยอมรับทางจิตใจว่าเป็นศูนย์ ในเนื้อเยื่อต่าง ๆ ศักยภาพของเมมเบรนจะมีลักษณะเฉพาะ ขนาดต่างกัน: สูงสุดในเนื้อเยื่อ m'yazovoi -80 -90 mV ในเนื้อเยื่อประสาท -70 mV ในความสุข -35 -40 mV ในเนื้อเยื่อเยื่อบุผิว -20 mV
Utvorennya MPP ที่จะนอนลงในความเข้มข้นของไอออน K + , Na + , Ca 2+ , Cl - และในลักษณะของเยื่อหุ้มของตาของคลิติน Zocrema ช่องไอออนซึ่งอยู่ในเมมเบรนอาจมีประสิทธิภาพ:
1. หัวกะทิ (เจาะเลือก)
2. ไฟฟ้า.
ในสภาวะที่สงบ ช่องโซเดียมจะปิด และช่องโพแทสเซียมส่วนใหญ่เปิดอยู่ ช่องสามารถม้วนงอและม้วนงอได้ ที่เมมเบรน ช่อง(ไม่เฉพาะเจาะจง) yakієเจาะองค์ประกอบทั้งหมด แต่เจาะทะลุโพแทสเซียมมากขึ้น ช่องโพแทสเซียมจะสั่นอยู่เสมอ และไอออนจะยุบตัวผ่านช่อง q ตามความเข้มข้นและการไล่ระดับเคมีไฟฟ้า
ตามทฤษฎีเมมเบรน-ไอออนิก การมีอยู่ของ MPP ถูกระบุ:
Ø โดยไม่มีการหยุดชะงักโดยการไหลเวียนของไอออนในช่องไอออนของเมมเบรน
Ø ความเข้มข้นของไอออนบวกที่เปลี่ยนแปลงอย่างต่อเนื่องทั้งสองด้านของเมมเบรน
Ø การทำงานของปั๊มโซเดียมโพแทสเซียมอย่างต่อเนื่อง
Ø การเจาะช่องต่างๆ สำหรับไอออนเหล่านี้
Іonіv K+ อุดมไปด้วย clitinum เสียงของโยคะมีขนาดเล็ก Na+ - ในทางกลับกันตำแหน่งนั้นอุดมไปด้วย clitinum และไม่เพียงพอใน clitinum Іonіv Сl – trochs ใหญ่กว่าชื่อของ clitin ตรงกลางล่าง เซลล์ตรงกลางอุดมไปด้วยแอนไอออนอินทรีย์ ซึ่งมีความสำคัญต่อการปกป้องประจุลบของพื้นผิวด้านในของเมมเบรน
ในเยื่อหุ้มเซลล์ที่สงบ มันถูกแทรกซึมโดยไอออน K+ เท่านั้น พวกเขามีโพแทสเซียมในสภาวะสงบ ระหว่างกลางความเข้มข้นสูงของ Na+ เมื่อคุณสงบลง พื้นผิวด้านนอกของเมมเบรนจะมีประจุบวก แอนไอออนอินทรีย์โมเลกุลสูง (โปรตีน) จะถูกทำให้เข้มข้นที่พื้นผิวด้านในของเมมเบรนและแสดงประจุลบ กลิ่นเหม็นที่เกิดจากไฟฟ้าสถิต utrimuyut K+ ไอออนจากด้านนอกของเมมเบรน บทบาทหลักของการส่องสว่าง MPP เป็นของ Do + ไอออน
หลังจากการไหลของไอออนผ่านช่องสัญญาณ ความแตกต่างในความเข้มข้นของไอออนจะไม่เปลี่ยนแปลง ดังนั้น ดูแลที่ไม่เปลี่ยนรูป ไม่มีอะไรที่เมมเบรนใช้ปั๊ม Na + - Up to + - กลิ่นเหม็นจะปล่อย Na+ ออกจากเซลล์อย่างต่อเนื่องและฉีดไล่ระดับความเข้มข้นที่ตรงกันข้ามเข้าไปใน K+ ไซโตพลาสซึม สำหรับ 3 Na + ion ซึ่งถูกพรากไปจากเซลล์จะมี 2 K + ion อยู่ตรงกลาง การถ่ายโอนไอออนเทียบกับการไล่ระดับความเข้มข้นเกิดจากการขนส่งแบบแอคทีฟ (จากพลังงานจำนวนมาก) ในช่วงเวลาที่มีพลังงานเอทีพี gyne
การแสดงศักยภาพของความสงบช่วยให้ลูกค้าบรรเทาลงได้จริงหลังจากแบ่งฝ่ายที่ตกอับเพื่อย้ายจากสภาวะสงบนิ่งไปสู่สภาวะตื่นตัว
เมื่อตื่นเต้น ขนาดของศักยภาพที่เหลือของเมมเบรนจะลดลงจากการชาร์จใหม่ ถ้าประจุภายในของเมมเบรนมีค่าเป็นลบน้อยลง แสดงว่ามีการสลับขั้วของเมมเบรนและเริ่มพัฒนาศักยภาพของได
4. ทริปโยคะ DIY และ mekhaizm ที่มีศักยภาพ
ระยะ Spivvіdshennia ของการตื่นขึ้นและระยะของdiїที่อาจเกิดขึ้น
ศักยภาพการตั้งชื่อศักยภาพของเยื่อหุ้มเซลล์ shvidke ซึ่งเป็นสาเหตุของการกระตุ้นเส้นประสาท m'yazovyh และ secretory clitin พื้นฐานของโยคะคือการเปลี่ยนแปลงการซึมผ่านของไอออนของเมมเบรน แอมพลิจูดและลักษณะของการเปลี่ยนแปลงในศักยภาพไม่เพียงพอที่จะอยู่ในความแรงของทีเซอร์ที่เรียกร้องมันเป็นสิ่งสำคัญมากขึ้นเพื่อให้ความแข็งแกร่งของ bula มีค่าไม่น้อยกว่าค่าวิกฤตเช่น เรียกว่าธรณีประตูแห่งการหยอกล้อ
เกณฑ์การหยุดชะงัก- แรงขั้นต่ำ tsya หากมีข้อผิดพลาดใด ๆ ปฏิกิริยาของแรงจะน้อยที่สุด เพื่อกำหนดลักษณะเกณฑ์ของ razdratuvannya vikoristovuyut เข้าใจ rheobase(Reo - ดีด, เบส - หลัก).
ครีมเกณฑ์แบ่งออก เกณฑ์ย่อย podrazniki, yakіสามารถเรียกร้องให้มีปฏิกิริยาในน้ำหรืออย่างอื่นเรียกร้องให้ทำลายการแลกเปลี่ยนคำพูดในคลิติน ดังนั้น suprathreshold โพดราซนิกิ
Vyniknuvshi, PD ขยายเมมเบรน ไม่เปลี่ยนแอมพลิจูดของมันเอง เฟสใหม่ถูกแยกออก:
1) การสลับขั้ว:
ก) ขั้วเต็ม;
b) การสลับขั้ว shvidka
2) การทำโพลาไรเซชัน:
ก) โพลาไรเซชันช้า;
b) การรีโพลาไรเซชันแบบเต็ม (ศักยภาพในการติดตามเชิงลบ)
3) Hyperpolarization (ศักยภาพการติดตามบวก)
คัดเลือก(เจาะไอออนได้เพียงชนิดเดียวเท่านั้น) เนื่องจากธรรมชาติของไอออน กลิ่นเหม็นทำให้ช่อง Na +, Ca ++, Cl-, K + ผ่านไปได้
ไม่คัดเลือก(เจาะไอออนหลายชนิด);
2) ตามวิธีการควบคุมจะแบ่งออกเป็น:
เงินฝากที่มีศักยภาพ(การกระตุ้นด้วยไฟฟ้า, potentiation)
ศักยภาพอิสระ(คีโมแอคทีฟ, (ลิแกนด์-รีเซพเตอร์-ฝาก), คีโมเคโรวานี)
mechanoexcitable(เครื่องจักรกล).
ศักยภาพในการทำให้จิตใจสงบ ทฤษฎีเมมเบรน-ไอออนนา ของศักยภาพเพื่อความอุ่นใจ Mіstseve i zbudzhennya กว้างขึ้น

เป็นที่ทราบกันดีอยู่แล้วว่าเมมเบรนของเซลล์ที่มีชีวิตนั้นมีโพลาไรซ์ พื้นผิวด้านในเป็นแบบอิเล็กโตรเนกาติเอทีฟสัมพันธ์กับสภาวะปกติ ศักยภาพของเมมเบรนเป็นสิ่งที่ดี – (ลบ) 70 – (90) mV เมื่อตื่นเต้น ขนาดของศักยภาพที่เหลือของเมมเบรนจะลดลงจากการชาร์จใหม่ การก่อตัวของศักยภาพการประหยัดนี้ถูกขัดขวางอย่างสงบโดยการไหลเวียนของไอออนอย่างต่อเนื่องตามช่องทางไอออนิกของเมมเบรน โดยความแตกต่างคงที่ในความเข้มข้นของไพเพอร์จากทั้งสองด้านของเมมเบรนโดยการทำงานอย่างต่อเนื่องของปั๊มโซเดียมโพแทสเซียม ความแตกต่างของความเข้มข้นของไอออนและโพลาไรเซชันของเมมเบรนจะคงอยู่ตลอดระยะเวลาการขับโซเดียมไอออนออกอย่างต่อเนื่องและการถ่ายโอนโพแทสเซียมไอออนไปยังเซลล์ ความเข้มข้นของโพแทสเซียมไอออนในเซลล์มีค่ามากกว่าความเข้มข้นของโพสต์คลิติน 30-40 เท่า ความเข้มข้นของโซเดียมหลังคลิตินจะสูงกว่าความเข้มข้นภายในคลิตินประมาณ 1 เท่า Електронегативність внутрішньої поверхні мембрани обумовлена наявністю в клітині надлишку аніонів органічних сполук, абсолютна величина потенціалу спокою (мембранний потенціал, трансмембранний потенціал, рівноважний калієвий потенціал) обумовлена головним чином співвідношенням внутрішньо- та позаклітинної концентрацій іонів калію та задовільно описується рівнянням เนิร์นสท:
 (1)
(1)
ทฤษฎีสมัยใหม่ของvrakhovuєเหมือนกัน:
1) ความแตกต่างของความเข้มข้นของโซเดียม คลอรีน แคลเซียมไอออน
2) การเจาะ (P) ของเมมเบรนสำหรับไอออนของผิวหนังต่อชั่วโมง
การแสดงศักยภาพของความสงบช่วยให้ลูกค้าบรรเทาลงได้จริงหลังจากแบ่งฝ่ายที่ตกอับเพื่อย้ายจากสภาวะสงบนิ่งไปสู่สภาวะตื่นตัว
Viniknennya ศักยภาพdії (depolarization)

ฉัน - ศักยภาพของเมมเบรน
1 - ศักยภาพในการพักผ่อน, 2 - การสลับขั้วเต็ม, 3 - การสลับขั้วช้า, 4 - การสลับขั้วช้า, 5 - การสลับขั้วเต็ม, 6 - การขั้วมากเกินไป
II - ความตื่นตัว
a - ปกติ, b - ขั้นสูง, - การหักเหของแสงแน่นอน,
d – การหักเหที่มองเห็นได้, e – ความเหนือปกติ,
e-subnormality
ศักยภาพdії (PD) พัฒนาขึ้นเนื่องจากการโพลาไรซ์ที่ชัดเจนของเมมเบรน (ความสงบที่อาจเกิดขึ้น) และเปลี่ยนการซึมผ่านของช่องไอออน (โซเดียมและโพแทสเซียม) ทันทีที่การเปลี่ยนแปลงที่อาจเกิดขึ้น การเปิดใช้งานช่องจะเพิ่มการเจาะสำหรับไอออน โซเดียมซึ่งเข้าสู่ไคลเอนต์และรับรองกระบวนการของการสลับขั้ว การรับเข้าสู่คลิตินของโซเดียมไอออนจะเปลี่ยนอิเล็กโตรเนกาติวีตี้ของพื้นผิวด้านในของเมมเบรน ซึ่งช่วยให้กระตุ้นช่องโซเดียมไอออนิกใหม่และการไหลเข้าของโซเดียมไอออนไปยังคลิตินเพิ่มเติม ความแข็งแกร่ง:
ก) แรงโน้มถ่วงของไฟฟ้าสถิตของกลุ่มประจุลบภายในเซลล์
b) การไล่ระดับความเข้มข้นของโซเดียมไอออน, ยืดตรงกลางเซลล์
จุดสูงสุดของศักยภาพของการเวียนหัวของประจุโซเดียมไอออนในคลิตินที่เท่ากันและในระยะทางที่เท่ากันภายใต้การไหลเข้าของแรงของอากาศในเวลาเดียวกันการชาร์จของไอออน
เยื่อหุ้มชีวภาพเป็นโครงสร้างที่ใช้งานได้จริงของคลิตินที่ล้อมรอบไซโตพลาสซึมและโครงสร้างคลิตินภายในส่วนใหญ่ สถาปนาสามัคคี ระบบคลิตินภายในท่อพับและปิดช่องว่าง
โครงสร้างพื้นฐานของเมมเบรนคือชั้นใต้ของฟอสโฟลิปิดซึ่งมีโปรตีนเมมเบรน ความหนาของเยื่อคลิตินคือ 6-12 นาโนเมตร โมเลกุลของไขมันมีลักษณะเป็นแอมโฟเทอริก ด้วยส่วนที่ชอบน้ำ กลิ่นเหม็นของสัตว์ร้ายอยู่ใกล้ตัวกลางน้ำ (เนื้อเยื่อระหว่างเซลล์และไซโตพลาสซึม) ส่วนที่ไม่ชอบน้ำของโมเลกุลจะถูกส่งไปยังตรงกลางของฟอสโฟลิปิด bisl โครงสร้างนี้เหมาะอย่างยิ่งสำหรับการแบ่งระยะหลังคลินิกและภายในคลินิก
โปรตีนที่เซลล์ขั้วของพวกมันฉีดเข้าไปในใต้บอลของฟอสโฟลิปิด จะสร้างพื้นผิวที่ชอบน้ำใกล้กับวัฏภาคของน้ำ เหม็น ฟังก์ชั่นต่างๆ: ตัวรับ เอนไซม์ ช่องไอออน ปั๊มเมมเบรน และตัวพาของไอออนและโมเลกุล
ความรู้ส่วนใหญ่ของเราเกี่ยวกับพลังของช่องไอออนเป็นผลมาจากการสร้างฟังก์ชันใหม่ Skin channel maє girl, ตัวกรองแบบเลือก, ประตูและกลไกของประตู keruvannya
ส่วนหนึ่งของช่องถูกสลักด้วยไฟฟ้า tobto จัดการการกระจายตัวของศักย์บนเมมเบรน (ช่องไอออนที่มีศักยภาพฝาก) สำหรับการสั่งซื้อครั้งแรกกับช่องสัญญาณจะมีเซ็นเซอร์ไฟฟ้าซึ่งขึ้นอยู่กับค่าของศักย์เมมเบรนหรือเปิดประตูของช่องหรือปิด
อีกทางเลือกหนึ่งสำหรับช่องไอออนิกคือเคราติไนเซชันของตัวรับ ประตูถูกควบคุมโดยโลคัสของตัวรับ ซึ่งกระจายอยู่บนพื้นผิวของเมมเบรน (เปิดเมื่อผู้ไกล่เกลี่ยโต้ตอบกับตัวรับ) ในช่องรับเซรามิกบางช่องระหว่างตัวรับและกลไกทั่วไป จะมีระยะกลาง (ประเภทแคมป์ตัวกลาง, โปรตีนไคเนส ฯลฯ)
แชนเนลเหล่านี้ให้พลังเมมเบรนที่สำคัญสองอย่าง: การคัดเลือกและการนำไฟฟ้า
หัวกะทิหรือความสั่นสะเทือนของช่องสัญญาณมั่นใจได้ด้วยโครงสร้างโปรตีนพิเศษซึ่งเป็นรูปทรงเรขาคณิตของช่อง
ตัวอย่างเช่น เส้นผ่านศูนย์กลางของโซเดียมไอออนคือ 0.19 นาโนเมตรพร้อมกับเปลือกไฮเดรตของไวน์ที่ใกล้เคียงกับ 0.3 นาโนเมตร คอของช่องโซเดียม 0.3 - 0.5 นาโนเมตร เพื่อที่จะผ่านเข้าไปในช่อง (โดยเฉพาะอย่างยิ่งผ่านตัวกรองแบบเลือกสรร) โซเดียมไอออนหรือไอออนอื่นมีความผิดในการอยู่ในรูปแบบของเมมเบรนไฮเดรตและเป็นไปได้น้อยที่บุคคลที่กำลังมองหา "เปล่า" จะผ่านช่อง ไอออนที่มากเกินไปไม่สามารถเข้าไปในคอได้ ไอออนที่เล็กเกินไปผิดธรรมชาติไม่สามารถนำเปลือกไฮเดรชั่นเข้าไปได้ ตัวกรองแบบเลือกออกจากช่องไม่ได้
ช่องโซเดียม (รูปที่ 6) ใช้เกท 2 ประเภท - การเปิดใช้งาน (m-gate) และการปิดใช้งาน (h-gate) ในใจของประตูการเปิดใช้งานที่สงบนั้นถูกปิด แต่เตรียมพร้อมที่จะบรรเทา และประตูการเปิดใช้งานก็พร้อม เมื่อ MP ลดลง (การสลับขั้วเป็น 60 mV) ประตูกระตุ้นจะเปิดขึ้นและปล่อยให้โซเดียมไอออนเข้าไปในคลิทินา จากนั้นประตูการปิดใช้งานจะเริ่มปิด (การทำให้ช่องโซเดียมไม่ทำงาน) หลังจากผ่านไปหนึ่งชั่วโมง ประตูการเปิดใช้งานจะเริ่มต้น ประตูการปิดใช้งานจะเปิดขึ้น และช่องสัญญาณก็พร้อมสำหรับรอบใหม่ ช่องนี้ถูกบล็อกโดย tetrodotoxin ยาชาเฉพาะที่ (novocain และใน)
ข้าว. 6. การทำงานของช่องโซเดียมและกลไก "เกตเวย์"
เอ - ที่ความสงบ m-gate ถูกปิด; B - เมื่อ zbudzhenny m-gate vіdkritі; B - ปิด h-vorit (ปิดใช้งาน) ระหว่างการสลับขั้ว
ช่องทางโพแทสเซียมยังสามารถเลือกได้ - สิ่งสำคัญคือต้องปล่อยให้โพแทสเซียมไอออนผ่านไป ถูกบล็อกโดยเตตระเอทิลโมเนียม กระบวนการระงับกลิ่นเหม็นจะแสดงออกมาอย่างอ่อน จากนั้นจะมีช่องโพแทสเซียมพิเศษที่เปิดใช้งานเมื่อความเข้มข้นของแคลเซียมภายในเซลล์เพิ่มขึ้นและการสลับขั้วของเยื่อหุ้มเซลล์เกิดขึ้น การเปิดใช้งานช่องที่มีโพแทสเซียม-แคลเซียมจะเร่งการรีโพลาไรเซชัน (MP ฟื้นฟู)
ช่องแคลเซียม แคลเซียมสตรัมขาเข้ามีขนาดไม่ใหญ่พอที่จะทำให้เกิดขั้วของเยื่อหุ้มคลิติน แคลเซียมทำหน้าที่เป็นตัวรอง
คนกลาง (ผู้ส่งสาร) การกระตุ้นของช่องแคลเซียมทำได้โดยการสลับขั้วของเยื่อหุ้มเซลล์ เช่น โดยกระแสน้ำโซเดียมเข้า การปิดใช้งานช่องแคลเซียมเกิดจากการเพิ่มขึ้นของความเข้มข้นภายในเซลล์ของแคลเซียมอิสระ โปรตีโอโปรตีนของไซโตพลาสซึมจับแคลเซียมซึ่งช่วยให้สามารถรักษาปริมาณแคลเซียมสตรูมาในปริมาณที่คงที่ได้หนึ่งชั่วโมงแม้ในระดับต่ำ ด้วยโซเดียมดีดนี้ จะเป็นการให้เกียรติมากขึ้น ช่องแคลเซียมถูกบล็อกโดยไอออนของแมงกานีส นิกเกิล แคดเมียม (2 วาเลนต์ไอออน) เช่นเดียวกับคำพูดทางการแพทย์ (verapamil)
แยกแยะระหว่างการขนส่งไอออนแบบพาสซีฟ (ปราศจากพลังงาน) และแอคทีฟ (การเก็บพลังงาน) ของไอออนผ่านเมมเบรน
เรื่อยไปสำหรับบัญชีของการแพร่กระจายที่เรียบง่ายและผ่อนคลาย
การแพร่กระจายอย่างง่ายเป็นไปตามกฎของฟิค - เบื้องหลังการไล่ระดับทางเคมี ไฟฟ้าเคมี หรือออสโมติก ตัวอย่างเช่น คลิตินมีโซเดียม 14 มิลลิโมลและ 140 มิลลิโมลตรงกลาง
สำหรับการแพร่กระจายง่าย ๆ แบบพาสซีฟ คำพูดอาจเป็นเรื่องอ้วน คำพูดที่ชอบน้ำไม่สามารถผ่านฟอสโฟลิปิดไบสเฟียร์ของเมมเบรนได้
การแพร่นั้นสะดวกทั้งเนื่องจากการมีช่องทางพิเศษหรือเนื่องจากการมีส่วนร่วมของพาหะ เนื่องจากพวกมันจับกับโมเลกุลที่ขนส่งโดยเฉพาะ จากนั้นจึงยึดตามการถ่ายโอนตามระดับความเข้มข้น
การขนส่งไอออนแบบแอคทีฟโดยปั๊มของเยื่อคลิตินช่วยรับรองการไล่ระดับไอออนิกจากทั้งสองด้านของเมมเบรน พลังงานสะท้อนให้เห็นในการเปลี่ยนคำพูดกับระดับความเข้มข้นของมัน
การมีส่วนร่วมในการขนส่งไอออนในระบบเอนไซม์พิเศษ - ATP-ases, yakіzdіysnyuyut hydrolysis ของ ATP แยก:
Sodium-potassium-ATP-ase ("ปั๊มโซเดียม") พบได้ในเยื่อหุ้มเซลล์ของ clitin ของสิ่งมีชีวิตทั้งหมด roslin และจุลินทรีย์ นี่คือโปรตีนเมมเบรนที่สามารถมีศูนย์รวมไอออนสองแห่ง หนึ่งในนั้น (โซเดียม) กระจายอยู่บนพื้นผิวด้านในของเยื่อหุ้มคลิตินส่วนอีกส่วนหนึ่ง (โพแทสเซียม) - บนพื้นผิวด้านนอก ตัวยับยั้งเฉพาะของเอนไซม์ є cardiac glycoside คือ strophanthin (ouabain) ซึ่งปิดกั้นปั๊มโซเดียม การไฮโดรไลซิสของโมเลกุล ATP หนึ่งโมเลกุลนั้นมาพร้อมกับการกำจัดโซเดียมไอออนสามตัวออกจากเซลล์และการฉีดโพแทสเซียมไอออนสองตัวโดยเซลล์ ด้วยการเพิ่มปริมาณโพแทสเซียมไอออนในเซลล์ตรงกลางหรือโซเดียมไอออนในเซลล์ตรงกลางของปั๊ม ปั๊มจะได้รับความช่วยเหลือ
Calcium-ATP-ase ("แคลเซียมปั๊ม") เป็นการแพร่กระจายที่กว้างที่สุดในเยื่อหุ้มเซลล์มะเร็งชนิด sarcoplasmic reticulum
Proton-ATP-ase ("โปรตอนปั๊ม") - ในเยื่อหุ้มของไมโตคอนเดรีย