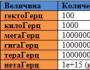
ulashtovaniの単純なウイルスのように。 ウイルスの構造。 ウイルスの分類の原則。 ウイルスの遺伝物質
Budovaとウイルスの分類
ウイルスが見られる 王国へビラ . ツェ
乾燥微生物(「ろ過する薬剤」)、
クリチンブドビ、バイオシンセティックシステムを洗わないでください、
悪臭は自律的な遺伝子構造であり、複製(複製)の特別な分離方法によって区別されます。細胞内で、ウイルスの核酸とそのタンパク質が合成され、次にウイルス粒子に折りたたまれます。
形成されたウイルス部分はと呼ばれます ビリオン。
ウイルスの形態と構造 hヘルプ 電子顕微鏡法, それらのサイズの破片は小さく、バクテリアの殻の質量と比較することができます。
vіrіonіvの形式は異なる場合がありますノイ(図):
指のような(Tyutyunのモザイクウイルス)、
kulepodіbnoi(ウイルスの話)、
球状(ポリオウイルス、VIL)、
糸のような(フィロウイルス)、
精子(豊富なバクテリオファージ)を探しています。
ウイルスの数は、次の要素によって決まります。
h 電子ヘルプ 顕微鏡学、
限外ろ過法による 目に見える細孔径のフィルターを通して、
方法 超遠心分離。
他の最大のウイルスはパルボウイルス(18 nm)とポリオウイルス(約20 nm)であり、最大のウイルスは天然ウイルス(約350 nm)です。
DNAとRNAを区別してウイルスを復讐するsi。悪臭を放つ 半数体、 Tobtomayut1セットの遺伝子。 Vinyatkomє二倍体ゲノムを含むレトロウイルス。 ウイルスのゲノムは、6から数百の遺伝子と表現から測定することができます 他の種類核酸:
ダブルスレッド、
シングルスレッド、
線形、
kіltsevymi、
断片化。
Єまた、陰性のRNAウイルス (マイナス鎖RNA)遺伝子議事録。 cixウイルスのRNAのマイナス文字列には、劣性機能しかありません。
別:
ただふわふわのウイルス (例えば、ポリオウイルス、A型肝炎)および
折りたたまれた折りたたまれたウイルス (たとえば、樹皮、インフルエンザ、ヘルペス、コロナウイルスのウイルス)。
で ウイルスの力だけ(米。) 核酸と呼ばれる白い殻で結ばれる キャプシド(Vidlat。 capsa- 場合)。 キャプシドは、繰り返される形態学的サブユニットで構成されています。 カプソメリック。核酸とキャプシドは、相互に1つずつ名前が付けられています。 ヌクレオカプシド。
で フォールディングウイルス(図)リポタンパク質排泄物のキャプシド シェル恥ずかしがり屋- スーパーキャプシド、カイペプロサム。 ウイルスの殻は、ウイルスに感染した細胞の膜の膜と同様の構造です。 obolontsiウイルスroztashovaniについて 糖タンパク質新着「ヒス」または「スパイク」 (ペプロミリ、また supercapsidniタンパク質)。 知られるべき特定のウイルスの殻の下 Mタンパク質。
そのような方法で、ただふわふわのウイルス 核酸とキャプシドで構成されています。折り畳み式のウイルス 核酸、キャプシド、リポタンパク質の殻で構成されています。
ビリオンマユット:
螺旋、
二十面体(キュービック)またはキャプシド(ヌクレオカプシド)に対する対称性の折りたたみタイプ。
スパイラルタイプヌクレオカプシドに対するギント様構造の絡み合いの対称性(例えば、インフルエンザウイルス、コロナウイルス)。 二十面体タイプウイルスの核酸(例えば、ヘルペスウイルス)に復讐するための、等尺性の空の体の溶液のキャプシドへの対称性。
キャプシドとシェル(スーパーキャプシド)は、過剰な培地からウイルスを保護し、歌うクリチンとの振動相互作用(吸着)、およびウイルスの抗原性と免疫原性に影響を与えます。
ウイルスの内部構造は 様 tsevinoy。 アデノウイルスでは、コアはDNAに結合したヒストン様タンパク質で構成されています。 reovirusiv-内部キャプシドのタンパク質から。
ウイルス学では、ウイルス学者はそのようなものを持っています それでソノミックカテゴリ :
苗字で終わる viridae),
podrodina(名前はで終わります virinae),
рід(名前はで終わる ウイルス).
ただし、すべてのウイルスに低値、特にピドロジンの名前が付けられているわけではありません。 バクテリアのように、二元的な名前を取り去ることのないウイルスの種。
ポロウイルスの分類の基礎次のカテゴリーの分隊:
核酸の種類vittya acidi(DNAまたは RNA)、 її構造、スレッドの数(1つまたは2つ)、特にウイルスゲノム開発のニュース(表2.3)、
ウイルスの拡大と形態、カプソメアの数と対称性のタイプヌクレオカプシド、シェルの存在(スーパーキャプシド)。
エーテルおよびデオキシコール酸に対する感受性、
クリッツの繁殖地、
抗原力と。
ウイルスは、とげのない生き物ととげのない生き物、そしてバクテリアと成長を攻撃します。 悪臭は人間の感染症の主な原因であり、発がんの過程にも関与しており、胎盤(風疹ウイルス、サイトメガ)などのさまざまな経路で感染する可能性があります。 lіїtaіn)、vrahayuchiは人々をplіdしました。 悪臭を放つことができます私を持ってきて感染後の状態-心筋炎、膵炎、免疫不全などの発症。
最も重要な(標準的な)ウイルスは感染性分子であり、これはウイルスであり、プリオンと呼ばれます。 プリオン- S. Prusinerによって造られた用語で、英語の単語「感染性の白い部分」のアナグラム。 正常なプリオンタンパク質(RgRS)のクリチン型は、人間を含む動物の生物に見られ、調節機能が低い。 それはPrP遺伝子をコードし、20番目のヒト染色体の短腕で腐敗します。 明らかに伝染性の海綿状脳症のプリオニー病(クロイツフェルト・ヤコブ病、クールー病など)では、プリオニータンパク質nabuvaєіnshoy、іnfektsіynoїの形 スクレイピー -きしみ、羊とキズのプリオン感染)。 cei感染性プリオンタンパク質はフィブリルのように見える場合があり、三次または四分の一構造を持つ通常のプリオンタンパク質のように見える場合があります。
ウイルスに近い他の目に見えないエージェント、є ウイロイド-環状のスーパーコイル状RNAの小分子
3.3。 ウイルスの生理学
ウイルス-内部クリチンの繁殖まではあまり一般的ではない、内部クリチン寄生虫を義務付けます。 ウイルスに感染した細胞では、さまざまな国でウイルスを転送することが可能です。
新しいビリオン番号の作成。
細胞の染色体から統合された状態でのウイルスへの核酸の移動(プロウイルスのように見える);
バクテリアのプラスミドを予測するカルシウム核酸としてのクリチンの細胞質の基礎。
したがって、ウイルスによって引き起こされる損傷の範囲も広い。細胞死で終わる顕著な生産的感染の場合、潜伏感染または悪性形質転換としてのウイルスとウイルスとの些細な相互依存に至る。
別 ウイルスとの3種類の相互作用ケージ付き:生産的、中途半端で統合的。
1. 生産型 - 新世代のビリオンの採用と感染細胞の死(溶解)(細胞溶解型)で終わります。 活動性のウイルスはクリチンから出てきますが、それらを破壊することはありません(非細胞溶解型)。
中途半端なタイプ - 新しいウイルスの採用が完了していないため、細胞内の感染プロセスの破片は1つの段階で中断されます。
統合型、 そうでなければ、ウイルス性は、クリチンの染色体におけるプロウイルスの存在下でのウイルスDNAの発達(統合)およびそのコンフルエンス(完全な複製)によって特徴付けられます。
ウイルスの繁殖(生産的)
生産的なタイプのvzaimodіїvіru saskіtina、tobto。 再生 ウイルス(lat。 再 - 繰り返し、 productio - virobnitstvo)、 6つの段階を経る:
1)吸着呼び出し中のビリオン;
2) 浸透クリチン中のウイルス;
3) 「razdyagannya」ウイルスゲノムのその非難(ウイルスの除タンパク);
4) 合成ウイルス成分;
5) 成形 virioniv;
6) virionivを終了しますクリチンから。
さまざまなウイルスでは、サイクルステージが逆になります
ウイルス吸着。ウイルス複製の最初の段階は吸着であり、ウイルスは細胞の表面に付着します。 2段階でリークします。 最初のフェーズは非特定です、 他のメカニズムを含む、ionnymtyazhennymizhウイルスおよびクリチナ。 別のフェーズ 吸着- ハイスペック チェスカ、 感受性クリチンの受容体の相同性、相補性に基づいて、ウイルスのタンパク質リガンドを「認識」します。 特定の細胞受容体を認識し、それらと相互作用するウイルスの表面上のタンパク質 、と呼ばれる 固定する tіlnimi タンパク質(主に糖タンパク質 іni)リポタンパク質シェルの倉庫で。
特定の受容体 クリチンは、タンパク質、脂質、タンパク質の炭水化物成分、脂質など、さまざまな性質を持つことができます。 したがって、インフルエンザウイルスの受容体は、dyhal経路の糖タンパク質および糖脂質(ガングリオシド)クレチンの貯蔵におけるシアル酸です。 Skazウイルスは神経組織のアセチルコリン受容体に吸着され、ヒト免疫不全ウイルスはTヘルパー細胞、単球、樹状細胞のCO4受容体に吸着されます。 1つの細胞には1万から10万の特定の受容体があり、数万から数百のウイルスがその細胞に吸着する可能性があります。
特定の受容体の存在は、歌う細胞、組織、器官のウイルス感染の活気の根底にあります。 ツェーそうランク 屈性 (ギリシャ語。 tropos - ターン、ターン)。 たとえば、肝臓細胞で最も重要に繁殖するウイルスは、肝向性と呼ばれ、神経細胞では神経向性、免疫担当細胞では免疫向性と呼ばれることがあります。
クリチンへのウイルスの侵入。ウイルスは、ウイルスのチュニックをクリチン膜で広げることによって、またはこれらのメカニズムの組み合わせの結果として、受容体休眠エンドサイトーシス(ビロペキシス)によってクリチンに侵入します。
1 . 受容体-休閑エンドサイトーシスクリチンによるビリオンの閉塞の結果として:ウイルスを復讐するために、ビリオンが付着したクリチン膜が閉じた内部クリチン液胞(エンドソーム)に埋め込まれます。 ATP沈着「プロトン」ポンプのチャンバーでは、エンドソームが酸性化され、エンドソーム膜を伴う折り畳まれたウイルスの怒ったリポタンパク質膜と、細胞のサイトゾルへのウイルスヌクレオカプシドの放出をもたらします。 エンドソームは、取り残されたウイルス成分を破壊するため、リソソームと結合します。 細胞質ゾルのエンドソームからエンベロープを持たない(単純に組織化された)ウイルスが出現するプロセスは、ほとんど発達していない。
2. クリチン膜を備えたZlittyaobtochuvannyaビリオン傷特定のチュニックウイルス(パラミクソウイルス、レトロウイルス、ヘルペスウイルス)にのみ典型的、 怒りの白。ウイルスタンパク質とクリチン膜の脂質との点相互作用があり、その結果、ウイルスリポタンパク質膜がクリチン膜と統合し、ウイルスの内部成分が細胞質ゾルに吸収されます。
しかし) ウイルスの「Rozdyagannya」(除タンパク)。その結果、内部コンポーネントの1つが振動し、建物が感染プロセスを引き起こします。 ウイルスの「発生」の最初の段階は、ウイルスと細胞膜の破壊によってクリチンに浸透する過程で、またはウイルスがエンドソームから細胞質ゾルに入るときに始まります。 ウイルスの「発生」の開始段階は、除タンパク部位への内部細胞輸送と密接に関連しています。 さまざまなウイルスについて、彼らはクリチンの「病気」の独自の専門分野を確立します。ピコルナウイルスの場合、細胞質では、リソソーム、ゴルジ装置が関与します。 ヘルペスウイルスの場合-海底核空間または核膜の細孔; アデノウイルスの場合-細胞質構造の半球、次にクリチンの核。 感染の最終産物は、核酸、核タンパク質(ヌクレオカプシド)、またはコアビリオンである可能性があります。 したがって、ピカルノウイルスの開発の最終産物は核酸であり、これは内部タンパク質の1つに共有結合しています。 そして、リッチシェルRNAウイルスでは、「破壊」の内因性産物はヌクレオカプシディまたはコアのいずれかであり、ウイルスゲノムの発現を変化させないだけでなく、さらに、細胞プロテアーゼの一部を保護し、さらなる生合成プロセスを調節します。
C)ウイルス成分の合成。ウイルスに対するタンパク質と核酸の合成、 なんて知識のバラそのスペースの時間に。 合成は細胞のさまざまな部分で行われるため、このウイルスの繁殖方法は dizジャンクティブ(Vidlat。 分離 --Roz'ednanny)。
と)ウイルスタンパク質の合成 . 感染した細胞では、ウイルスゲノムは2つのグループのタンパク質の合成をコードします。
1. 非構造タンパク質、さまざまなヨガの段階でウイルスの体内繁殖にどのように役立つか。
2. 構造タンパク質、ウイルスウェアハウスに入る方法(ゲノム、ウイルスゲノム、キャプシドおよびスーパーキャプシドタンパク質に関連)。
前非構造的なbіl カム 1)ウイルスゲノムの転写と複製を確実にするRNAまたはDNAの合成のための酵素(RNAまたはDNAポリメラーゼ)。 2)調節タンパク質; 3)構造タンパク質の強いパターンによる不安定性の影響を受けるウイルスタンパク質の前線。 4)修飾する酵素 ウイルスタンパク質例えば、プロテイナーゼおよびプロテインキナーゼ。
タンパク質の合成クライアントの場所で、適切なプロセスを実行することが賢明です 転写 (Vidlat。 転写 - 遺伝子情報を核酸から情報RNA(iRNA)のヌクレオチド配列に「書き換える」ことにより) 放送(Vidlat。 翻訳 - 伝達)-タンパク質のバックボーンからのリボソーム上のiRNAの読み取り。 ウイルスの異なるグループでのiRNA合成中の崩壊情報の伝達は同じではありません。
私 . DNAウイルスは同じように遺伝子情報を実現します。 スキームによると、細胞ゲノムのように:
ゲノムDNAウイルス-»文字起こしiRNA-" ブロードキャストウイルスへのタンパク質。
さらに、DNAいたずらウイルスはこのプロセスのために振動しますクリチンポリメラーゼ(ウイルス、そのゲノムは細胞の核で転写されます-アデノウイルス、パポウイルス、ヘルペスウイルス)またはRNAポリメラーゼ(ウイルス、そのゲノムは細胞質で転写されますなど)。 )。
II 。 プラス鎖RNAウイルス (たとえば、ピコルナウイルス、フラビウイルス、そして gavirusi)ゲノムを刈る、vikonuєとは iRNA機能; vinrozpіnaєsyaであり、リボソームによって翻訳されています。 これらのウイルスのタンパク質の合成は、スキームに従って転写作用なしで実行されます。
ゲノム RNAウイルス->タンパク質のウイルスへの翻訳 .
III. マイナス一本鎖RNAミストのゲノム ウイルス (オルトミクソウイルス、パラミクソウイルス、ラブドウイルス)および双子(レオウイルス)は、核酸をウイルスに結合するRNAポリメラーゼの関与により、iRNAを転写するためのマトリックスとして機能します。 それらのタンパク質の合成は、次のスキームに従います。
ゲノム RNAウイルス-»文字起こしta- RNA- ブロードキャスト ウイルスへのタンパク質。
IV. レトロウイルス (ヒト免疫不全ウイルス、発癌性レトロウイルス)は、遺伝子情報を転送する独自の方法を持っている可能性があります。 レトロウイルスのゲノムは、二倍体である2つの同一のRNA分子で構成されています。 レトロウイルスの倉庫には、特定のウイルス特異的酵素(ベルターゼ転写酵素またはレベルターゼ)があり、その助けを借りて病原性転写のプロセスが実行されるため、一本鎖相補DNA(cDNA)がゲノムRNAマトリックス上で合成されます。 DNAの相補鎖は二本鎖相補DNAからコピーされるため、細胞ゲノムに組み込まれ、細胞DNAが枯渇したRNAポリメラーゼの助けを借りてiRNAに転写されます。 これらのウイルスのタンパク質の合成は、次のスキームに従います。
ゲノム RNAウイルス->補完 DNA-»文字起こし iRNA
-"ブロードキャスト ウイルスへのタンパク質。
ウイルスゲノムの複製 すなわち、ウイルスが折りたたまれたときに活発である外部ウイルスゲノムのコピーのクリチンにおける蓄積をもたらすウイルス核酸の合成。 ゲノムを複製する能力は、ウイルス中の核酸の種類、ウイルス特異的または細胞性ポリメラーゼの存在、およびウイルスの性質に依存し、クリチンでのポリメラーゼの確立を誘導します。
複製のメカニズムはウイルスによって異なり、次のような場合があります。
1)二重DNA;
2)一本鎖DNA;
3)プラス一本鎖RNA;
4)マイナス一本鎖RNA;
5)二本鎖RNA;
6)同一のプラス鎖RNA(レトロウイルス)。
1. ツインLNKウイルス . 二本鎖ウイルスDNAの複製は、独特で保存されたメカニズムによって観察されます。DNA鎖が絡み合った後、それらに相補的な新しい鎖が得られます。 新しく合成された皮膚DNA分子は、1つの同じ合成された鎖で構成されています。 これらのウイルスの前には、線形のdvunitkovu DNA(たとえば、ヘルペスウイルス、アデノウイルス、ポックスウイルス)、またはパピローマウイルスのようなキルトセビのようなウイルスの大きなグループがあります。 ポックスウイルスを含むすべてのウイルスでは、ウイルスゲノムの転写は核で起こります。
複製のユニークなメカニズムは、ヘパドナウイルス(B型肝炎ウイルス)の特徴です。 ヘパドナウイルスゲノムは二本鎖環状DNAで表され、一方の鎖はもう一方の鎖の短い(非線形プラス鎖)です。 穂軸は収穫されます(図3.7)。 次に、再び、細胞のDNA沈着RNAポリメラーゼの助けを借りた二本鎖DNAは、iRNAの小分子と完全に一本鎖のプラスRNAの導入によって転写されます。 残りはプレゲノムRNAと呼ばれます。 ウイルスゲノム複製のテンプレートとして受賞。 iRNAの合成は、ウイルスRNA側副DNAポリメラーゼ(逆転写酵素)を含むタンパク質の翻訳プロセスに関与します。 細胞質に移動するこの酵素の助けを借りて、プレゲノムRNAは転写されてDNAの負の鎖に戻るため、それ自体がDNAのプラス鎖を合成するためのテンプレートとして機能します。 このプロセスは、間違ったプラス鎖のDNAに復讐するために、二本鎖DNAの除去で終了します。
一本鎖DNAウイルス . 一本鎖DNAウイルスの唯一の代表はパルボウイルスです。 双子のウイルスゲノムを作成するための活発な細胞DNAポリメラーゼのパルボウイルス、いわゆる複製型の残り。 この場合、負のDNA鎖(プラス鎖)は負のDNA鎖上で相補的に合成され、新しいビリオンのプラスDNA鎖の合成のテンプレートとして機能します。 並行して、iRNAが合成され、ウイルスペプチドの翻訳が行われます。
プラス一本鎖RNAウイルス . サイウイルスには、ピコルナウイルス、フラビウイルス、トガウイルスなどのウイルスの大きなグループが含まれ(図3.8)、RNAのゲノムプラス鎖がiRNAの機能を無効にします。 たとえば、クリチナに侵入した後、ポリオウイルスRNAはリボソームと結合してiRNAのように機能し、これに基づいて優れたポリペプチドが合成され、RNA枯渇RNAポリメラーゼ、ウイルスプロテアーゼ、キャプシドタンパク質の断片に分割されます。 ゲノムプラス鎖RNAに基づくポリメラーゼはマイナス鎖RNAを合成します。 timchasovysubvіynaRNAが形成され、それは中間複製ランカと呼ばれます。 中間の複製スリランカは、繰り返されるRNAのプラス鎖と、数値的に頻繁に完成するマイナスストリングで構成されています。 すべてのマイナス鎖が確立されている場合、悪臭は、RNAの新しいプラス鎖を合成するためのテンプレートのように振動します。 このメカニズムは、ウイルスへのゲノムRNAの複製、および多数のウイルスタンパク質の合成として勝利を収めています。
マイナス一本鎖RNAウイルス。 マイナス一本鎖RNAウイルス(ラブドウイルス、パラミクソウイルス、オルソミコウイルス)は、RNAが沈着したRNAポリメラーゼを倉庫に保管します。 クリチナに浸透したゲノムマイナス鎖RNAは、ウイルスRNA-側副RNA-ポリメラーゼによってRNAの上下のプラス鎖に変換されます。 不完全なコピーは、ウイルスタンパク質の合成におけるiRNAの役割を示しています。 子孫ゲノムRNAのマイナス鎖を合成するためのテンプレートの新しいコピー(中間段階)
ツインRNAウイルス。 これらのウイルス(レオウイルスおよびロタウイルス-rus)の複製のメカニズムは、マイナス一本鎖RNAウイルスの複製に似ています。 転写の過程で、プラス鎖がiRNAとして機能し、複製に関与することは明らかです。つまり、RNAのマイナス鎖を合成するためのテンプレートとしてです。 RNAのプラス鎖との複合体に残って、ビリオンゲノム二本鎖RNAが隠されています。 これらのウイルスのウイルス核酸の複製は、細胞の細胞質で起こります。
6 . レトロウイルス (プラススレッドの二倍体RNAウイルス)。 レトロウイルス逆転写酵素は、(RNAウイルステンプレート上で)マイナス鎖のDNAを合成します。これは、プラス鎖のDNAを採用されたサブ鎖のDNAにコピーし、環状部で閉じます(図3.10)。 DNAのDalísubvіyna鎖は染色体細胞、utvoryuyuchiプロウイルスです。 多数のビリオンRNAは、クリチンDNA沈着RNAポリメラーゼの関与により、組み込まれたDNAの鎖の1つが転写された結果として定着します。
成形ウイルス。ウイルスは自己折り畳み経路によって形成されます。ウイルスの倉庫部分は、ウイルスの選択場所である核の細胞または細胞の細胞質に輸送されます。 Z'ednannyakomponentіvvirionuobumovリノ疎水性、イオン性、水性結合および立体的生存率の存在。
Іsnuyutそう慈善の原則 ウイルスの選択 :
ウイルスの成形は、ポリペプチドの倉庫の背後にある成熟したウイルスの形で開発された、承認された中間形態による多段階プロセスです。
単純に爆破されたウイルスの保管ウイルス核酸とキャプシドタンパク質および確立されたヌクレオキャプシドとの相互作用における極。
折りたたみ式武装ウイルスヌクレオカプシドは背面に形成され、クリチン膜(おそらくウイルスのリポタンパク質エンベロープ)によって相互に修飾されます。
さらに、クリチナムの核で複製するウイルスの選択は核膜の関与によって影響を受け、細胞質で複製するウイルスの選択は小胞体膜または原形質膜の膜の関与によって影響を受ける。 、糖タンパク質が吸収される場所。
折りたたまれたvlashtovanihの数マイナス鎖RNAウイルス(オルトミクソウイルス、パラミクソウイルス)のウイルスは、修飾されたクリチン膜の派生物であるマトリックスタンパク質(Mタンパク質)のタイトルを持っています。 マユチの疎水性優勢は、ヌクレオカプシドとウイルスのリポタンパク質膜の間の仲介役の役割を勝ち取ります。
□ 折り畳み式のウイルス成形プロセスでは、脂質や炭水化物など、定規の細胞の有効成分を倉庫に含めます。
Vіhіdvіrusіvіzkіtini。ウイルス複製の次のサイクルは、5〜6年(インフルエンザウイルスなど)またはsprat deb(ヘパトウイルス、皮質ウイルスなど)で完了します。 ウイルスの繁殖のプロセスは、細胞からの出口で終わります。これは、vibukhovymパスまたはbrunkuvannyam、エキソサイトーシスです。
Vibukhovyの方法: klіtiniのgynaeから、多数のvirionivsが一度に出てきます。 vibukhovyパスでは、リポタンパク質の殻を洗うように、ウイルスの細胞から簡単に抜け出すことができます。
Nirkuvannya、exocshpt リポタンパク質膜の洗浄などのウイルスの力、ヤク єクリチン膜に似ています。裏面のヌクレオカプシドは沈降します。そうでない場合、ビリオンのコアは細胞膜に輸送され、そこでウイルス特異的タンパク質がすでに導入されています。 次に、ヌクレオカプシドまたはビリオンのコアがクリチン膜と接触する領域で、これらの細胞の成長が始まります。 形成されたニルカは、使い古されたウイルスのように見えるクリートのように見えます。 このクリチナを使用すると、長期的な命を救い、ウイルスの子孫を生み出すことが可能です。
細胞質で形成されるウイルスは、原形質膜(例えば、パラミクソウイルス、トガウイルス)、または細胞の表面への遠い出口を有する小胞体膜の膜(例えば、ブニヤウイルス)のいずれかを貫通することができる。
細胞の核で形成されるウイルス(例えば、ヘルペスウイルス)は、改変された核膜を通って核周囲空間に侵入し、リポタンパク質膜をそのようなランクで満たす。 次に、悪臭は細胞質小胞の倉庫から細胞の表面に運ばれます。
ウイルス-他のすべての生物。その範囲は20〜300nmです。 平均的な悪臭では、バクテリアは50個少なくなります。 光学顕微鏡の助けを借りて、バクテリアを通過させないように悪臭がフィルターを通過するのを助けることはできません。
ウイルス アウトブレイク
後継者はしばしば食べ物について疑問に思います、なぜ彼らは生きているのですか ウイルス? 生体構造を尊重し、それが遺伝物質(DNAまたはRNA)であるかどうかにかかわらず、それを自己生成するために作成する場合、それは固体でなければなりません。つまり、ウイルスは生きています。 生きていることのしるしとして、クリチンの生命の存在を尊重してください、それは否定的です:ウイルスは生きていません。 次に、ウイルスのクリチン定規の姿勢が自己作成に知られていないことを付け加えます。
再発見のために ウイルスについて進化の過程で何が起こっているのかを知る必要があります。 ウイルスが遺伝物質であること、原核細胞と真核細胞の「vt_k」を取得し、それが提出された瞬間まで建物を取得することを許可する必要があります。クリチン、それは研ぎ澄まされています。
ウイルスのクリチンによるポーズただし、完全に不活性な状態であるためには、クリチナiに再浸透し、それらを命令に従属させ、それらと同一のコピーを多数作成するために必要な一連の命令(遺伝暗号)を持っている場合があります(ウイルス)。 次に、ウイルスの進化の過程が後でクリチンに現れたことを認めることは論理的です。

ブドバウイルス
ブドバウイルスさらに簡単です。 悪臭は前進する構造から形成されます:
1)コア-DNAまたはRNAのいずれかで表される遺伝物質。 DNAまたはRNAは片面または両面にすることができます。
2)capeida-コアをしみ出させるzahisnoyタンパク質シェル。
3)ヌクレオカプシド-コアとキャプシドで構成される折り畳み構造。
4)シェル-VILやインフルエンザなどのウイルスのように、宿主細胞の原形質膜に似た付属肢のリポタンパク質ボールを持っている可能性があります。
5)カプソメア-カプソメアがしばしば誘導される、繰り返される同一のサブユニット。

キャプシドの空洞の形は、高度な対称性によって突き刺されています、 ウイルスの建物結晶化するまで。 Tseを使用すると、X線結晶学の方法と追加の電子顕微鏡の両方でそれを行うことができます。 ウイルスのサブユニットがklitz-mastersに確立されるとすぐに、悪臭はすぐに単一のウイルス粒子に統合されます。 ウイルスの存続期間のスキームが簡略化され、小さなものが示されています。
構造物の場合 ウイルスキャプシド典型的な歌唱タイプと対称性、特に多面体とスパイラル。 多面体はバガトヘドロンです。 ウイルスの多面体は最も広く、20の三角形の面、12のカット、30のリブを持つ二十面体です。 赤ちゃんAには通常の二十面体があり、赤ちゃんBにはヘルペスウイルスがあり、その粒子の中に162個のカプソメリックが二十面体に組織化されています。

らせん対称性の最後の図は、小さなものの証拠かもしれません、 RNAウイルス tyutyunovyモザイク(TMV)2130個の同一タンパク質カプソメアのthウイルスのキャプシド。
tm buv 最初のウイルス、すっきりと見えます。 ウイルスに感染すると、病んでいる植物の葉に黄色いひよこが現れます-これは葉のモザイクの名前です(図2.18、B)。 ウイルスrozpovsyudzhuyutsyaは、病気が成長した場合、または一部の部分が健康なroslinsでdotikに到達した場合、またはタバコからの煙でpovitryanymの方法で、そのようなbulovikoristanoに感染した葉の準備のために任意または機械的に発生します。
ウイルス、バクテリアを攻撃し、グループを確立するので、それらはバクテリオファージまたは単にファージと呼ばれます。 一部のバクテリオファージは、はっきりとはっきりとした二十面体の頭と尾を持ち、らせん対称です)。 小さなものを狙うのは概略です 活動的なウイルスの画像それは彼らの世界観を示しています
私たちの惑星の豊かな歴史の中で、すべての動植物の発達の近くで、目に見えない警備員が絶えず導入されました。ウイルス(ラテンウイルス-otruta)。
ウイルスの微視的なロズミールに関連して、生物のような折りたたみ可能な内部の豊かなクリチンの生活が緩和されたので、悪臭は生きているクリチンにとってははるかに少なく、細菌にとってはより豊かです。 私は、人、生き物、爬虫類、肋骨だけでなく、さまざまな植物を含むすべての生物にウイルスを注ぎ込んでいます。
電子顕微鏡のワイン造りの後、20世紀の穂軸でのみ、未亡人は貧しい医療従事者の目に揺れることができました。それについては、その瞬間まで、すでに非人格的な理論がありました。 ウイルスの歌と人々は、その世界の形の背後で彼らの間で震えました。 病気の一種として休閑していると、さまざまな病気の症状がさまざまな形で現れます。皮膚のやけど、内臓、流木などです。
ウイルス感染
1852年、ドミトリー・ヨシポビッチ・イワノフスキー(ロシアの植物学者)は、モザイク病の一種であるroslintyutyunから感染性抽出物を取り除くことができました。 この構造は、Tyutyunovのモザイクウイルスの名前を取り除いた。
ウイルスへのブドバ
 ウイルス部分の中心にはゲノムがあります(DNAまたはRNA構造で表されるため情報が減少します-位置1)。 ゲノムの周りで、タンパク質シェルによって表されるキャプシド(位置2)が拡張されます。 キャプシドのタンパク質シェルの表面には、リポタンパク質シェルがあります(位置3)。 真ん中のシェルは波立たせられたcapsomeryです(位置4)。 革のカプソメアは、1本または2本の白の糸で構成されています。 皮膚ウイルスのカプソメアの数は、ほぼ一定です。 カプソメアの数をその数のカプソメアに復讐する皮膚ウイルス 他の種類ウイルス
ウイルス部分の中心にはゲノムがあります(DNAまたはRNA構造で表されるため情報が減少します-位置1)。 ゲノムの周りで、タンパク質シェルによって表されるキャプシド(位置2)が拡張されます。 キャプシドのタンパク質シェルの表面には、リポタンパク質シェルがあります(位置3)。 真ん中のシェルは波立たせられたcapsomeryです(位置4)。 革のカプソメアは、1本または2本の白の糸で構成されています。 皮膚ウイルスのカプソメアの数は、ほぼ一定です。 カプソメアの数をその数のカプソメアに復讐する皮膚ウイルス 他の種類ウイルス  敢えて言うなら。 Deyakіウイルスはそれ自身のタンパク質シェル(キャプシド)には見られません。 このようなウイルスは単純と呼ばれます。 まず第一に、ウイルスは、自分たちの生活のように、もう1つの層(追加のリポタンパク質)の殻を持っています。これはフォールディングと呼ばれます。 ウイルスには2つの生命体があります。 ウイルスのPozaklіtinnazhittєva形式は呼ばれます ヴァリオン(落ち着いて、ochіkuvannya)。 ウイルスが活発に繁殖するため、ウイルスの内部形態は植物性と呼ばれます。
敢えて言うなら。 Deyakіウイルスはそれ自身のタンパク質シェル(キャプシド)には見られません。 このようなウイルスは単純と呼ばれます。 まず第一に、ウイルスは、自分たちの生活のように、もう1つの層(追加のリポタンパク質)の殻を持っています。これはフォールディングと呼ばれます。 ウイルスには2つの生命体があります。 ウイルスのPozaklіtinnazhittєva形式は呼ばれます ヴァリオン(落ち着いて、ochіkuvannya)。 ウイルスが活発に繁殖するため、ウイルスの内部形態は植物性と呼ばれます。
ウイルスの優位性
Віруси не мають клітинної будови, їх відносять до найдрібніших живих організмів, відтворюються всередині клітин, мають просту будову, більшість із них викликають різні хвороби, кожен тип вірусу розпізнає та інфікує лише певні типи клітин, містять лише один тип нуклеїнової кислоти (ДНК або РНК) 。 。
ウイルスの分類

体のクリチンがスピーチを征服するように
他の生物に基づいて、子孫を作るためのウイルスは生きた細胞を必要とします。 自分でワインを増やすことはできません。 たとえば、人類の細胞は核で構成されています(新しいDNAのDNAは遺伝子地図であり、細胞の計画は生命を支えることです)。 Ядро клітини оточує цитоплазма, в якій розташовані мітохондрії (вони виробляють енергію для хімічних реакцій, лізосоми (у них розщеплюються матеріали, що надійшли з поза), полісоми і рибосоми (в них виробляються білки і ферменти для здійснення хімічних реакцій, що відбуваються в клітці) .Klitini細胞質、バーニッシュは、それに慣れるために、そして同じように見えるように、最高の運河に浸透します。細胞質で腐敗するように、中心への命令。
ウイルスの繁殖
 もっと意図されていたように、ウイルスは似たような細胞を作るために生きた細胞を必要とします。それはそれ自体では繁殖できないからです。 ウイルスがクリチンに侵入するプロセスは、いくつかの段階で構成されています。
もっと意図されていたように、ウイルスは似たような細胞を作るために生きた細胞を必要とします。それはそれ自体では繁殖できないからです。 ウイルスがクリチンに侵入するプロセスは、いくつかの段階で構成されています。
ウイルスが細胞に侵入する最初の段階は、細胞の表面(標的)の沈殿物(電気的相互作用による吸着)ヨガです。 クリティーナは、母親の表面的な受容体の心臓を非難するメタです。 実行可能な表面受容体がなければ、ウイルスは細胞に到達できません。 したがって、電気的相互作用の結果としてドアにやってきたそのようなウイルスは、strushuvannyaパスでクリーンアップすることができます。 ウイルスがクリチナに侵入する別の段階は、不可逆性と呼ばれます。 受容体が存在するため、ウイルスはクリチナムとタンパク質の棘に付着するか、糸がクリチナムの受容体と相互作用し始めます。 細胞受容体は、皮膚ウイルスに特異的なタンパク質または糖タンパク質として機能します。
第3段階の1時間以内に、ウイルスは濡れます(移動します) 細胞膜細胞内膜フグの助けを借りて。
第4段階では、クリチン酵素がウイルスタンパク質を分解します。このようにして、ウイルスゲノムとの「リンク」から発生し、DNAおよびRNA構造で表されるため、崩壊情報が分解されます。 次に、RNAヘリックスは急速に拡大し、細胞の核に向かいます。 細胞の核では、ウイルスのゲノムが変化しています 遺伝情報クライアントと自分自身を実現します。 ロボットのそのような変化の結果として、細胞は無秩序になり、細胞内の必要なタンパク質と酵素の置換は、ウイルス(修飾された)タンパク質と酵素を合成し始めます。
 ウイルスがクリチナに侵入してから新しい亜種がリリースされるまでに経過した時間は、到着と呼ばれ、潜伏期間と呼ばれます。 それは時々(vіspa、インフルエンザ)から数日(kіr、アデノウイルス)に変わる可能性があります。
ウイルスがクリチナに侵入してから新しい亜種がリリースされるまでに経過した時間は、到着と呼ばれ、潜伏期間と呼ばれます。 それは時々(vіspa、インフルエンザ)から数日(kіr、アデノウイルス)に変わる可能性があります。
悪臭のきしみ:表面、水、土壌、および物体の表面。 フローリングの悪臭は小さいので、標準的な顕微鏡ですべてのタイプを見ることができるわけではありません。 Tseウイルス、不思議な自然の偉業、戦争の終わりまでyakіはなく、razyuchuvizhivannyaかもしれません。
知っている:otruynyと危険
ウイルスは、ラテン語からヨガを翻訳するかのように、その名前に完全に忠実です:otruta。 以前は、この単語は解析せずにすべての医療従事者に慣れていました。 しかし、19世紀になると状況は変わりました。
200年前、特定の病気の影響を受けたtyutyunの葉を使った実験の過程でのロシアのイワノフスキーの教えは、水と細菌のvmistの追加フィルターのためにジュースが追加されたとしても、すべての生体材料が健康を救うことを発見しました。 彼らは、口蹄疫ウイルスや黄熱病など、ろ過の方法で新しいタイプの攻撃的な薬剤を見始めました。 「フィルタリング」という段階的な言葉が登場し、科学の発展のこの段階では、全世界でより多くの病気を求める人々は通常ウイルスと呼ばれています。
生きていても死んでもいない
Tsepitanyadosiє科学的なsuperechokの主題。 右側では、その時間に、まるで未来のウイルスが醸造されたかのように(私たちにとっては、Tyutyunovのモザイクを呼び出して)、彼らの行動計画、そして彼らが考えているかのように重要な詳細が明らかにされました:私は生きています、私は死んでいる、どうしたの?

引数:
- 分子構造;
- ゲノムに復讐する。
- 気候の真ん中は積極的に搾乳する必要があります。
反対意見:
- 空の女性の姿勢は絶対に不活性です。
- 自分でタンパク質を合成しないでください。クリチンロードなしで遺伝物質を配布することはできません。
構造的特徴
多くの病気を引き起こすブドバウイルスは、詳細について疑問に思いますが、よくできた米がたくさんある可能性があります。 Nasampered、ウイルスの原因となる形態はビリオンと呼ばれます。 ワインは次の要素で構成されています。
- 1〜3分子の核酸を含む核;
- キャプシド-脾臓から酸を保護するタンパク質の袋。
- タンパク質-脂質層から形成される殻(実際には開始しないでください)。

核酸はウイルスの遺伝暗号です。 Tsikavo、scho nicoli deoxyribonucleic acid リボ核酸同時に迷子にならないでください。 その時間に、微生物のように、疑いの余地のない「活気」の中で、例えばクラミジアは、彼らの倉庫に不快な酸を持っているかもしれません。 遺伝子情報に関しては、1〜3個の遺伝子で囲むことができますが、そうでない場合は、最大100個の遺伝子にすることができます。
ビリオンの補遺殻を占有生物に配置し、芽の細胞に変化を加えた。 補足できるウイルスは、細胞質核膜を凍結し、その断片から二次嚢を形成することができます。 さらに、そのような力の殻は、ヘルペスやインフルエンザウイルスなどの優れた標本とは同等ではありません。
ビリオンの構成要素は、保護の機能として機能し、情報を収集し、それらの必要な突然変異のウイルス複製に責任があります。
フォームウイルス
ウイルスの存在の特徴は、キャプシドの形でそれらの分類を横たえるようなものです。

一番 ただのウイルスキャプシドの倉庫にある種のタンパク質分子が存在しているように見えるので、そうかもしれません。 これは裸のウイルスの名前であるため、シェルからきれいに解放されます。
カプソメアで覆われたAleєvirioni-単純な幾何学的形状を構成するほんの数個の分子。 Budovaウイルス、およびそれらのカプソメアに名前を付けると、攻撃的なエージェントの識別に重要な役割を果たします。 形はかなり異なります:尾のある頭、ストレートカット(vispa)、袋(flu)、スティック(Tyutyunのモザイク)、糸(ジャガイモの球根の病気)、バガトヘドロン(polyomielitis)、kulepod_bny( skaz)。
ナノスペース
ウイルスは小さいので、電子顕微鏡でしか詳しく見ることができません。 形態とウイルスが同じでなければ、バクテリアは常により大きなサイズ(約50倍)で成長します。 ビリオンのサイズは、小さい(20-30 nm)から大きい(400 nm)までの範囲で変化します。

クリティーナの職業
クリチナへのウイルスの侵入は同じ順序で屈服しません-本質的に、同様のメカニズムは他のどこでも発生しません。 クリチナビリオンのポーズは、眠っている結晶化したスタンによって変化しています。 活発な子供たちが生まれ変わるので、エール・ヴァルトは空のバガンでヨマを食べます。
- 吸着。言い換えれば、フレーム化された細胞の壁へのウイルス(150)の付着。
- Viropexis。プロットを介してウイルスに付着している、クリチンでの非中間のzanurennyaのプロセス。 Tsikavyの瞬間:クリチナは侵入を通過せず、ウイルスのその部分に、virnishe、yogoタンパク質は、クリチナによって「私たち自身」として識別されます。
- 畳語。ウイルスがクリチンで増殖した場合にのみ、感染性の侵入が始まります。 それらは、それら自体の分子と同様に、数値キャプシドを満たす新しい分子を合成します。
- 撤退。 過密の瞬間、居間は破壊され、ウイルスは何も流れず、悪臭は新しい細胞に対して振動しています。 どのような状況でも、このようなプロセスはデキルコムの方法で実行できます。
それは素晴らしいことですが、微生物はクリチンにとって何百倍も少なく、ロボットを破壊するのは簡単で、交換プロセスに破壊的に押し込み、しばしば犠牲者を破壊します。
ウイルス侵入の種類
それは、クリチナス破壊の性質と攻撃的なエージェントの些細なことのために落ちる分類に似ています。 cymに関連する感染には3つのタイプがあります。
- 破滅的:どのタイプの感染が溶解性と呼ばれるか、新しいウイルスの場合、それはクライアントの空間から大規模に実行可能であり、独自の方法ですべてを破壊し、新しいクリチンを征服しようとします。
- 安定、または永続的:細胞の働きを破壊することなく、名前のウイルス塊の段階的な活性化を特徴とする;
- 添付ファイル:潜伏型は、クリチン染色体のウイルスゲノムの発達中に発達し、その後、クリチンが分離されると、ウイルスを娘構造に伝達します。

ヴァルトの終わりには、これらの微視的な物質の敵対的な変動が示されています。これが、警戒されている症状の違いの理由です。 DNA(ヘルペス、ビスパ)の存在下でウイルスを調査し、RNA(バクテリオファージのスプラット)である口蹄疫を排除します。 Krіmіnshhogo、іvіrіoni復讐lipіdi。
その他のオプション:脂質を含まないウイルス。アデノウイルスと同様に、バクテリオファージの大部分がより重要です。
それは、世界の人生の早い段階で、生命の形態を秩序づけ、人々の欲望にそれらをもたらすことを学ぶであろうその状況を奨励します。
ウイルス。 ウイルスの分類。 Tipivzaєmodіїkіtinivіrusіv
Rozmіri-15〜2000 nm(deyakіroslinウイルス)。 ウイルスの生き物と人間の最大の平均は、450nmまでの自然の生命労働者です。
ごめん ウイルスが殻を洗う- キャプシド、より少ないタンパク質サブユニットで構成されています( capsomiriv)。 より多くのウイルスのカプソメアは、らせん状および立方対称を作成できます。 Vіrіonizіspiralsіmеrієyumаyutpalichkopodіbnuフォーム。 スパイラルタイプの対称性の背後では、より多くのウイルスが誘導され、増殖は敵対的です。 より多くのウイルス、 敵対的な細胞人や生き物は、立方体の対称性を作り出すことができます。
折りたたみ
折りたたみ ウイルスは、宿主の細胞の原形質膜の一部である糖タンパク質を含むリポタンパク質表面膜によってさらに覆われる可能性があります(たとえば、ウイルスウイルス、B型肝炎)。 スーパーキャプシド。 糖タンパク質の助けを借りて、主の細胞の壁の表面にある特定の受容体の認識とそれにウイルス部分の付着が観察されます。 炭水化物では、糖タンパク質の細胞が先のとがった棒のようにウイルスの表面から突き出ています。 dodatkovaチュニックは、細胞コリのウイルス部分の場所への浸透が吸収されるため、細胞の原形質膜に腹を立てることがあります。 追加のシェルには、宿主細胞でのウイルス核酸の合成やその他の反応を確実にする酵素を含めることができます。
バクテリオファージは折りたたまれたブドフを搾乳することができます。 Їxはフォールディングウイルスに持ち込まれる可能性があります。 たとえば、T4バクテリオファージは、頭、つぼみ、尾の糸などの拡張部分で構成されています。 頭部は、核酸を含むキャプシドに折りたたまれています。 つるには、コミッサール、空のせん断、収縮し、推測で伸びたバネを備えたシースによる離乳、および尾のスパイクと糸を備えた基部プレートが含まれています。
ウイルスの分類
ウイルスの対称性、外殻の存在の存在におけるフィールドでのウイルスの分類。
| デオキシウイルス | リボウイルス | ||
| DNA dvolantsyuzhkova | DNA odnolantsyuzhkova | RNA dvolantsyuzhkova | RNA odnolantsyuzhkova |
| 対称の立方型: –楕円形の膜なし(アデノウイルス); -羊の殻(ヘルペス)から | 対称の立方型: –外膜なし(死んだファージ) | 対称の立方型: –外殻なし(レトロウイルス、ロスリン牧場パフのウイルス) | 対称の立方型: –羊の膜なし(エンテロウイルス、ポリオウイルス) らせん対称型: -ovnishnіh膜なし(Tyutyunのモザイクウイルス); -Zovnishnimiシェル(インフルエンザ、skaz、発癌性RNAウイルス) |
| 対称性のタイプの変更(Tペアバクテリオファージ) | |||
| 対称性のタイプを歌わずに(vispi) | |||
ウイルスの生存率は、生物の細胞ではそれほど顕著ではありません。 核酸は、ゴスポダールのクリチンのウイルス粒子の合成に関与しています。 ウイルスの姿勢は生命の兆候を示さず、 ビリオン .
ウイルスのライフサイクルは、次の2つのフェーズで構成されます。 pozaklitinny(ビリオン)、yakіyのvinは活力の兆候を示していません、それ 内部クリチン 。 体のウイルス部分は感染前に1時間も費やしません。 たとえば、ポリオウイルスは、感染活動を数か月間、長期間保存することができます。 B型肝炎ウイルスは短時間の煮沸のために保存されます。
一部のウイルスの活発なプロセスは、核、他のウイルス(細胞質、他のウイルス)、核、および細胞質で発生します。
Tipivzaєmodіїkіtinivіrusіv
いくつかのタイプを含む、細胞とウイルスの間の相互作用:
- 生産的 -核酸は、新世代の声の合成のマスターの細胞にウイルスを誘発します。
- 中途半端 -繁殖はどの段階でも中断され、新世代は落ち着きません。
- ウイルス性 -ウイルスの核酸は宿主細胞のゲノムに導入され、複製することはできません。