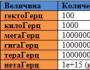
受容体機能とは何ですか? 細胞膜の受容体機能。 クリティーニ受容体:理解、roztashuvannya、rіznovidi、budova
グリコカリックス-膜内在性タンパク質-糖タンパク質の多糖類ランスを復讐するリポタンパク質膜ボールに関連する因果関係。
原形質膜の最も重要な機能の1つは、タンパク質または糖タンパク質の性質を持つ可能性のある膜に存在する受容体装置の助けを借りて、細胞を安全に通信(接続)することです。 原形質膜の受容体溶液の主な機能は、音信号の認識であり、細胞の静脈は正しく配向され、組織は分化の過程で確立されます。 さまざまな調節システムの活動は、受容体機能に関連しており、免疫応答を形成します。
主要部分:
そのため、細胞表面の受容体は、膜タンパク質または糖タンパク質の要素(糖タンパク質)を突き出すことができます。 スピーチの音に非常に敏感であるため、農民は細胞の表面に広がるか、小さな領域で拾うことができます。
生物の異なる細胞は、異なる受容体のセット、または1つの同じ受容体の異なる感度を持つことができます。
豊富な細胞受容体の役割は、特定の発話の伝達と物理的作用物質に応答する能力、および細胞の表面から細胞の中央への細胞間信号の伝達に影響を及ぼします。 この時間で、活性ホルモンの助けを借りて細胞に信号を送信するシステムが十分に開発され、ペプチドランスが入る倉庫に送られます。 ホルモンは、細胞の中央に浸透することなく、システムの細胞質の受容体部分と特異的に相互作用し、アデニル酸シクラーゼ(原形質膜の細胞質部分に近いタンパク質)を活性化して、サイクリックAMPを合成します。 残りは、内部の細胞酵素または酵素のグループを活性化または抑制します。 このようにして、コマンド(原形質膜からの信号)がセルの中央に送信されます。 アデニル酸シクラーゼシステムの効率はすでに高いです。 したがって、ホルモンの1つまたは複数の分子の相互作用により、非人格的なサイクリックAMP分子が合成され、信号が1000倍に増加する可能性があります。 で この特定のタイプに変換音信号として機能するアデニル酸シクラーゼシステム。
細胞の表面にある受容体のセットの多様性と特異性は、マーカーの複雑なシステムの作成につながります。これにより、見知らぬ人から自分の細胞(個人または同じ種)を調べることができます。 同様の細胞が相互依存して1つずつ入り、表面に粘着性が生じます(最も単純な細菌の接合、組織細胞複合体の確立)。 一連の決定因子マーカーの影響を受ける異なる細胞の場合、それらはそれらを受け入れないか、そのような相互作用でスイッチが切られるか、または他の生物では免疫反応の結果として減少します。
特定の受容体の局在は、物理的作用物質がどのように反応するかである原形質膜に関連しています。 そのため、原形質膜、または同様の光合成細菌や藍藻類では、受容体タンパク質(クロロフィル)が局在しており、光量子と相互作用します。 光受容体タンパク質(ロドプシン)の特別なシステムが、生物の感光性細胞の原形質膜で開発されました。これにより、光信号が化学信号に変換され、電気インパルスが生成されます。
原形質膜を通過する能動輸送を見る
短い:
- 一次能動輸送は輸送ATPアーゼによって制御されるため、イオンポンプの名前が付けられました。
- 二次能動輸送-能動輸送の過程で作成された、第2の音声の濃度の勾配の追加のエネルギーで、第3の濃度のプロ勾配の音声の膜を介して転送します。
最近:
能動輸送は、輸送アデノシントリホスファターゼ(ATPアーゼ)によって媒介され、ATP加水分解のエネルギーに依存しています。
スピーチの能動輸送を参照してください:
- 一次能動輸送、
- 二次能動輸送。
一次能動輸送
低濃度の媒体から高濃度の媒体への音声の転送は、大まかな勾配では説明できません。 拡散。 このプロセスは、ATPの加水分解のエネルギーまたはエネルギーに基づいており、任意のイオン、特にすべてのナトリウムの濃度勾配があります。 音声の能動輸送のためのエネルギー源として、それはATPの加水分解であり、膜を通る他の分子やイオンの動きではなく、輸送は主に能動と呼ばれます。
主にアクティブな転送はトランスポートATP-aseによって実行されるため、イオンポンプの名前が削除されました。 生き物の細胞では、Na +、K + --ATPase(ナトリウムポンプ)が最も広く拡張されています。 内在性タンパク質原形質膜Ca2+-ATPase。これは、小胞体(小胞体)の原形質膜にあります。 3つのタンパク質はすべて、リン酸化して酵素の中間リン酸化型を確立する能力という大きな力を持っている可能性があります。 リン酸化された植物では、酵素は2つのコンフォメーションで変化する可能性があり、通常はE1とE2と呼ばれます。 酵素コンフォメーションは、最初の分子のポリペプチドランスの空間配向(パッキング)の方法です。 酵素の2つの指定されたコンフォメーションは、イオンに対する胞子形成性を変化させることを特徴としています。 輸送されているイオンを表示するためのさまざまな建物。
二次能動輸送
二次能動輸送は、能動輸送の過程で生成される、第2の音声の濃度勾配のエネルギーの助けを借りて、第2の濃度のプログラジエントの音声膜を介した移動です。 動物では、二次能動輸送の主なエネルギー源は、Na + /K+-ATPaseのために作成されたナトリウムイオン濃度勾配のエネルギーです。 たとえば、小腸の粘膜のクリチン膜には、上皮細胞におけるNa +グルコサイトの移動(共輸送)に寄与するタンパク質が含まれています。 その場合、指定されたタンパク質からのグルコースとすぐに結合するNa +が電気化学的勾配によって輸送されるため、グルコースの輸送が発生する可能性は低くなります。 電気化学的Na+勾配は、細胞からのこれらの陽イオンの能動輸送によって刺激されます。
ロボットのNa+ポンプの脳では、メディエーターの中毒(再吸収)と関連しています。これは、めまいの要因の場合に神経終末に見られる生理学的に活発な発話です。
心筋細胞およびNa+の機能を有する平滑細胞において、K +-ATPase結合Ca2+輸送は原形質膜を介して、細胞タンパク質の膜にタンパク質が存在し、Na+およびCa2+タンパク質輸送(対向輸送)を促進します。 それらは、ナトリウムイオンおよびナトリウムイオンの濃度勾配のエネルギーと引き換えに、クリチン膜に移されます。
クリチンでは、内部の細胞プロトンであるNa + /H+交換体で酸化イオンをナトリウムに交換するタンパク質が明らかになりました。 ceiトランスポーターは、細胞内pHの安定性を維持する役割を果たします。 Na + /Ca2+およびNa+/H+が影響を受ける流動性-膜を通過するNa+の電気化学的勾配に比例する交換。 Na +のクリチン後濃度の変化、強心配糖体によるNa +、K + -ATPaseの阻害、またはカリウムを含まない培地では、カルシウムとプロトンの濃度が増加します。 Na +、K +-ATPaseの阻害中にCa2+の内部細胞濃度を増加させる目的は、心拍数を強化するための強心配糖体の臨床診療におけるうっ血の基礎です。
クリチン膜に局在し、発話の伝達のメカニズムのように、異なる輸送ATPアーゼは、分子付着の主要な要素であるポンプであり、振動粘土を提供し、クリチンによる歌われた発話(例えば、電気リチウム)のポンピングを提供します。 非電解質の能動的特異的輸送(分子輸送)は、ポンプやキャリアなど、さまざまな種類の分子機械の助けを借りて実装されています。 非電解質(単糖、アミノ酸、その他のモノマー)の輸送は、シンポート(他の音声の輸送)を使用して実行できます。その流れは、最初のプロセスの濃度勾配とエネルギーの背後にあります。 Symportは、ATPを使用せずに、イオン勾配(ナトリウムなど)で処理できます。
輸送ATPaseは、ATPをエネルギーで分解する高分子輸送タンパク質です。 このプロセスは、能動輸送の背後にある原動力です。 このようにして、プロトンが移動します(プロトンポンプまたは無機イオン(イオンポンプ))。
能動輸送は、エンドサイトーシスおよびエキソサイトーシスによって阻害されます。
エンドサイトーシス-粘土化した硬い粒子(食作用)または発話(飲作用)による原形質膜の浸潤による球根の採用。 tsomaのせいにされている滑らかなまたはoblyamovanny球根は、ファゴソームまたはピノソームと呼ばれます。 卵子のエンドサイトーシスの方法は、黄色のタンパク質を変色させること、白血球が異物や免疫グロブリンを変色させること、ニルコビ細管が一次セクションからタンパク質を吸収することです。
エキソサイトーシスは、エンドサイトーシスに似たプロセスです。 ゴルジ装置からのさまざまな球根、リソソームが原形質膜から注ぎ出され、名前が変わります。 この場合、plashechkaの膜は原形質膜に芽を出すか、またはmichurの形で細胞質に変わることができます。
Zahisna関数
タンパク質は、微生物を殺して追い出すのを助けることができるので、血液や他の故郷で発見されます。 血漿の倉庫に抗体(タンパク質、他の外来物質と同じ種類の微生物を認識する皮膚)が入り、補体系のタンパク質の分泌を見つけます。 2番目のクラスの抗体(ci-タンパク質は免疫グロブリンとも呼ばれます)で、最も広いものは免疫グロブリンGです。リゾチームタンパク質は、ムレインと破滅的な細菌壁を分解する酵素です。 細胞がウイルスに感染すると、生物はインターフェロンタンパク質を認識します。インターフェロンタンパク質はウイルスを複製し、新しいウイルス粒子を生成します。
微生物毒素のようなタンパク質-コレラ毒素、ボツリヌス毒素、ジフテリア毒素など。私たちの体のPoshkozhuyushchy細胞は、悪臭が微生物を私たちから保護します。
受容体機能
タンパク質は信号を送受信するのに役立ちます。 生理学では、細胞受容体の理解はありません。 歌う信号を受信する細胞(たとえば、細胞には受容体があります)。 クリチン受容体のエール、受容体タンパク質がロボットに追加されます。 したがって、目に位置するタンパク質ロドプシンは、光の量をキャプチャし、その後、ポディアのカスケードが細胞の細胞で始まり、神経インパルスの終了と信号の伝達につながります脳。
受容体タンパク質は、受容体クリチンだけでなく、他のクリチン受容体にも含まれています。 体内でさらに重要な役割を果たしているのはホルモンです。スピーチは一部のクリチンに見られ、他のクリチンの機能を調節します。 ホルモンは、クリチン標的の表面または中央にあるホルモン受容体などの特殊なタンパク質と結合します。
規制機能
タンパク質を含む豊富な(すべてではありませんが)ホルモン-たとえば、下垂体および視床下部のすべてのホルモン、インスリンおよびその他
多くの白人は、機能の散りばめに勝つことができます。
タンパク質高分子はb-アミノ酸で構成されています。 多糖類の倉庫がその「1つ」(1つと2つ)に入り始めるとすぐに、それは何度も繰り返され、タンパク質は20の異なるアミノ酸から合成されます。 さらに、ザイブランタンパク質の分子として、タンパク質の倉庫にあるデュースアミノ酸残基は化学変化を検出できるため、「成人」タンパク質では30を超える異なるアミノ酸残基を検出することができます。 これが、安全性のためのモノマーの多様性と、タンパク質に関連する生物学的機能の多様性です。
b-アミノ酸は次のようになります:

ここでR-異なるアミノ酸の異なる原子グループ(ラジカル)。 カルボキシル基に最も近い炭素原子はギリシャ語の文字bで示され、b-アミノ酸の分子の閉じたアミノ基のすぐ次の原子です。
中性媒体では、アミノ基は弱い塩基性力とH +イオンの付加を示し、カルボン酸基はイオンに対してわずかに酸性で解離性の結合を示すため、分子の総電荷は一度に変化しませんが、私たちはすぐに正と負に帯電したグループを運びます。
休耕地では、ラジカルRの性質上、疎水性(非極性)、親水性(極性)、酸性、および月のアミノ酸が区別されます。
酸性アミノ酸には別のカルボキシル基があります。 光学酸のカルボキシル基の場合は3倍強力です。アスパラギン酸では、カルボキシルの半分がpH 3.86で解離し、グルタミン酸ではpH 4.25で、オクトボイン酸では4.8でしか解離しません。 アミノ酸の中で最も強いのはアルギニンです。鎖状ラジカルの半分はpH11.5で正電荷を帯びます。 リシンでは、鎖ラジカルが典型的な一級アミンであり、vinはpH9.4で半分イオン化されています。 月のアミノ酸の中で最も弱いのはヒスチジンであり、そのイミダゾール環はpH6で半分プロトン化されています。
親水性(極性)のものの中には、生理学的pHでイオン化する2つのアミノ酸もあります-システイン、いくつかのSHグループでは、H +イオンは、シルボドニュ、チロシンと同様に、いくつかの弱酸性フェノールグループで追加できます。 健康の保護はさらに弱い悪臭を放ちます:pH 7では、システインのイオン化は8%、チロシンは0.01%です。
b-アミノ酸の検出には、バイコリストのニンドリン反応を使用します。アミノ酸がニンドリンと相互作用すると、青色の生成物がはっきりと生成されます。 さらに、他のアミノ酸は独自の特定の反応を示します。 したがって、芳香族アミノ酸は硝酸で醸造する必要があります(反応時間中に芳香環が追加されます)。 培地を使用すると、感染はオレンジ色に変化します(感染の変化と同様に、メチルオレンジなどの指標に変わります)。 キサントプロテインの名前でのこの反応は、タンパク質の検出にも活発であり、タンパク質の大部分の破片は芳香族アミノ酸です。 ゼラチンは反応を示さず、石はチロシン、フェニルアラニン、またはトリプトファンに復讐しない可能性があります。 プライムナトリウムNa2PbO2と一緒に加熱すると、システインは硫化鉛PbSの黒色沈殿物を溶解します。
Roslinsと豊富な微生物は、単純な無機スピーチからアミノ酸を合成できます。 クリーチャーは数個以上のアミノ酸を合成し、残りを取り除くことができます。 このようなアミノ酸は必須と呼ばれます。 人間にとって必須のものは、フェニルアラニン、トリプトファン、スレオニン、メチオニン、リジン、ロイシン、イソロイシン、ヒスチジン、バリン、アルギニンです。 穀物文化がリジンやトリプトファンとほとんど関係がないのは残念です。 より多くの量マメ科植物のmіstjatsya。 農民の非暴力的な伝統的な子供たちは、穀物やマメ科植物のように聞こえます。小麦(zhito)とエンドウ豆、米と大豆、トウモロコシと豆は、さまざまな大陸でのこのような人々の台頭の典型的な尻です。
20アミノ酸すべての炭素のb原子は、sp3混成サイトにあります。 4つのリンクはすべて109°に近いため、アミノ酸の式は四面体で記述できます。
perekonatisyaは簡単です、schoは1つの鏡像のように2種類のアミノ酸にすることができます。 広場で動かして振り返っても、まとめることはできません。悪臭は右手と左手のように分かれています。
この種の異性体は光学異性体と呼ばれます。 中央の原子は反対側に炭素(vinは非対称中心と呼ばれます)を持っているので、Vinはその場合にのみ可能です(そのため、グリシンには光学異性体がありませんが、19個のアミノ酸があります)。 アミノ酸の2つの異なる異性体から、図1に示すもの。 1は右利き用のフリルで、D字型と呼ばれ、zlivaはL字型と呼ばれます。
基本的な物理 化学力アミノ酸のD-異性体とL-異性体は同じで、屈折力が異なります。それらの違いは、反対側の光の偏光面を包み込みます。 これらの反応の違いは、他の光学活性物質に関連しています。
人間へのウイルスに含まれるすべての生物のタンパク質の倉庫に入るのはL-アミノ酸だけであることは明らかです。 D型は、真菌や細菌によって合成されるいくつかの抗生物質に見られます。 タンパク質は、その倉庫に複数のタイプのアミノ酸異性体が含まれているため、その場合にのみ規則正しい構造を形成できます。
重要な役割は、細胞の活力と膜の受容体機能によって果たされます。 それは、化学的および物理的要因の特定の認識に関連する、特別な構造(受容体タンパク質)の原形質膜への局在化に関連しています。 悪臭の悪臭の外側から、多くの浸透性タンパク質と糖タンパク質が、槍の多糖類フレークに復讐します。 分子アンテナのクリチン「喪失」をカバーするこれらの糖タンパク質のいくつかは、ホルモン受容体の役割を果たします。 歌うホルモンがその受容体と結合すると、糖タンパク質の構造が変化し、クリチン反応が始まります。 スピーチの歌がクリチンで見つかるか、それから派生する必要があるなど、チャネルが開かれます。 クリチン表面には、さまざまな薬剤との特定の反応を可能にする受容体の大きなセットが含まれている場合があります。 豊富な細胞受容体の役割は、細胞内のシグナル伝達呼び出しの伝達に関与しています。
22. Clitini受容体:理解、roztashuvannya、rіznovidi、budova。
細胞の原形質膜には、受容体の名前をとったシグナル分子であるタンパク質が広がっていた。 クリチン受容体は分子に結合し、作用を開始します。 悪臭は膜貫通タンパク質によって表され、ホルモンや神経伝達物質などの生理学的に活性な分子を結合するための特別な空間を作り出します。 sing分子の結合における豊富な受容体タンパク質は、膜の輸送力を変化させます。 その結果、膜の極性が変化したり、神経インパルスが発生したり、発話の交換が変化したりする可能性があります。
細胞内受容体と原形質膜近くの細胞の表面にある受容体を認識します。 それらの中で、2つのタイプの受容体が見られます-クリチニチスのチャネルに結合することとチャネルに結合しないことです。 悪臭は、歌うターゲットの信号の速度と活気を求めて互いに戦うことです。 化学受容体(ホルモン、神経伝達物質)との相互作用に続いて、チャネルに接続された受容体は、開いたチャネルの膜に付着し、その後、透過性が即座に変化します。 チャネルに関連しない受容体も化学受容体と相互作用しますが、性質が異なり、最も重要なのは酵素です。 ここでは、部族ではありますが、その効果は間接的で、明らかに刺激的です。 これらの受容体の機能は、その記憶の形成の根底にあります。
23.クリチンシェルを介したスピーチの伝達:理解、rіznovidi、お尻。
膜輸送-スピーチをクリチン膜を越えてクリチンに、またはクリチンから輸送します。これは、単純拡散、促進拡散、能動輸送などのさまざまなメカニズムによってサポートされています。 さまざまなタイプの輸送が16と17のタイプで説明されています。
24.異文化間の接触:理解、rіznovidi、意味。
クリチン間接触-クリチン間で作られ、タンパク質の助けを借りて確立されました。 悪臭は、クリチンの間に中間の呼び出しがないことを確認します。 さらに、クリチニーは、クリチナス間スピーチを介して送信される追加の信号(ヘッドランク-シグナルスピーチ)のスタンドで1つずつ混ざり合っています。
皮膚タイプの細胞間接触は、特定のタンパク質のラフノクに対して形成され、そのような数が多いほど- 膜貫通タンパク質。 特別なアダプタータンパク質は、細胞骨格との細胞間接触のタンパク質を橋渡しすることができ、特別な「骨格」タンパク質は、折り畳み式の超分子構造でこれらのタンパク質の分子の周りを橋渡しすることができます。 豊かな窪地では、Ca2 +イオンが増加すると、間質の筋が崩壊します。
th受容体。 体の真ん中で、ホルモン、神経伝達物質、または代替品のいずれかを呼び出します。これは、医薬品およびアルコール(アゴニスト)としてzastosovuyutsyaです。 Deyakіligandi、navpaki、ブロック受容体(拮抗薬)。 器官の感覚、発話のリガンドがある場合は、受容体に香りを注入するか、味わってください。 さらに、経口受容体の分子は光に反応し、聴覚器官や点状器官では、受容体は刺し傷や他のサブかみそりによって呼ばれる機械的注入(圧力または張力)に敏感です。 膜電位の変化に反応する感熱性受容体タンパク質と受容体タンパク質もあります。
受容体のクラス[ | ]
細胞受容体は、膜受容体と細胞内受容体の2つの主要なクラスに分けることができます。
膜受容体[ | ]
「アンテナ」の機能は、音声信号の認識です。 Dіlyankidvohsusіdnіhkіtin、shchorazznіyutは、クリチンの接続を保護し、1対1でリンクすることができます。 Zavdyaki tsymu clitinyは方向付けられ、分化の過程で組織を作成します。 認識可能な細胞は、さまざまな方法で知られているように、さまざまな分子で存在し、なぜ悪臭は、認識される相補的な細胞である可能性があるクリチンによって振動的に粘土化されます(たとえば、LPGはLDL受容体の助けを借りて崇拝されています)。
膜受容体の2つの主要なクラスは、代謝型受容体とイオノトロピック受容体です。
イオノトロピック受容体は、リガンドに結合すると開閉する膜チャネルです。 Ionni Strumi、TsomoのVinikayat、ViclicayuZmіni膜貫通RIZNITSIPotzіalivI、vnaslіdokTsoyo、Zbudinosti Klitini、そして彼は内部濃縮物のヘビであり、彼らは陰謀のシステムの評価に生き残ることができます。 最近開発されたイオノトロピック受容体の1つは、n-コリン作動性受容体です。
から形成されるGタンパク質の構造 3種類単位(ヘテロ三量体)-αt/αi(黒)、β(赤)およびγ(緑)
代謝型受容体は、内部細胞メディエーターのシステムによって結合されます。 リガンドと結合したときにそれらのコンフォメーションを変化させて、生化学反応のカスケードを開始させ、おそらく細胞の機能状態を変化させます。 膜受容体の主な種類:
- ヘテロ三量体Gタンパク質に結合する受容体(たとえば、バソプレシン受容体)。
- 内因性のチロシンキナーゼ活性を示す受容体(例えば、インスリン受容体または上皮成長因子受容体)。
Gタンパク質、7つの膜貫通ドメイン、N末端および細胞内C末端を含む膜貫通タンパク質にリンクされた受容体。 リガンドとの結合部位は亜クリチナスループ上にあり、Gタンパク質との結合ドメインは細胞質のC末端近くにあります。
受容体の活性化は、αサブユニットがβγサブユニット複合体に解離して活性化される前に実行されます。 もしそうなら、それは酵素を活性化するか、さもなければ不活性化し、二次中間体を生成します。
チロシンキナーゼ活性を持つ受容体は、多くの場合プロテインキナーゼによっても、発症した細胞内タンパク質をリン酸化し、このようにしてシグナルをクリチンの内部に伝達します。 構造によると、それらは1つの膜ドメインを持つ膜貫通タンパク質です。 原則として、ホモ二量体、そのようなコーティングのサブユニットは、ジスルフィドパッチで覆われています。
細胞内受容体[ | ]
細胞内受容体-原則として、転写因子(例えば、糖質コルチコイド受容体)または転写因子と相互作用するタンパク質。 内部細胞受容体のほとんどは、細胞質内のリガンドに結合し、活性状態に移行し、リガンドと一緒に細胞核に輸送され、そこでDNAに結合し、特定の遺伝子または遺伝子のグループの発現を誘導または抑制します。
一酸化窒素(NO)の形成のための特別なメカニズム。 膜を透過するこのホルモンは、細胞質ゾルのグアニル酸シクラーゼ、一酸化窒素受容体、および二次中間体であるcGMPを合成する酵素に結合します。
ホルモンシグナルの細胞内伝達の主なシステム[ | ]
アデニル酸シクラーゼシステム[ | ]
アデニル酸シクラーゼシステムの中心部分は、ATPからcAMPへの変換を触媒する酵素アデニル酸シクラーゼです。 この酵素は、G sタンパク質(英語刺激下)によって刺激されるか、G iタンパク質(英語抑制下)によって無視されます。 次に、cAMPはcAMPが枯渇したプロテインキナーゼ(プロテインキナーゼA、PKAとも呼ばれる)に結合します。 タンパク質エフェクターのその遠いリン酸化の活性化を引き起こす必要があり、それはクリチンにおいて生理学的役割を果たすでしょう。
ホスホリパーゼ-カルシウム系[ | ]
G qタンパク質は酵素ホスホリパーゼCを活性化し、PIP2(膜ホスホイノシトール)をイノシトール-3-リン酸(IP3)とジアシルグリセリドの2つの分子に分割します。 これらの分子の皮膚は2番目の中間体です。 IP3は、小胞体の膜上の受容体に結合することができます。これにより、カルシウムが細胞質に放出され、さまざまな細胞反応が開始されます。
グアニル酸シクラーゼシステム[ | ]
このシステムの中心分子はグアニル酸シクラーゼであり、GTPからcGMPへの変換を触媒します。 cGMPは、低酵素とイオンチャネルの活性を調節します。 グアニル酸シクラーゼのІsnuєスプラットアイソフォーム。 それらの1つは、心房性ナトリウム利尿受容体に間接的に結合している一酸化窒素NOによって活性化されます。
cGMP制御交換は、腸と腸でのイオン輸送につながり、心臓ではリラックスのシグナルとして機能します。
受容体の薬理学[ | ]
原則として、健康な受容体は、主要な内因性リガンドだけでなく、他の構造的に類似した分子にも結合します。 この事実により、受容体と結合し、顔や拭き取りなどの状態を変化させる、vikoristovuvatyの外因性発話が可能になります。
受容体機能は、細胞の最も重要な機能であり、外部および内部環境の信号に適切に応答し、それによって細胞が心の心にとどまることができます。
シグナル--tserіznіスピーチまたは他の方法でクリチンの情報を送信するエネルギーを参照してください。 信号は次のようになります。
化学物質-ホルモン、メディエーター、成長因子、サイトカインなど。 臭いのスピーチ、さもなければ彼らは味わいを醸し出します。
物理的-光、音、温度、圧力、電位;
物理化学的-浸透圧バイス、圧力Pro2またはCO2;
折り畳み。
クリチン受容体-遺伝的に決定された高分子。さまざまな属の細胞に局在し、化学的および物理的性質の生物学的に重要な特定の信号の受信に特化しています。 構造の背後にある受容体は、3つのドメインで構成されています。
1)膜の後ろ-信号音声との安全な接続-リガンド;
2)膜貫通-信号を転送し、変換前に構築します。
3)細胞質-内部細胞プロセスの確保-シグナルへの応答。
クリチン受容体は2つのグループに分けられます:
原形質膜受容体;
細胞内受容体は細胞質と核です。
原形質膜受容体プラスモレミウムおよび組織の表面へのプレーティングは、リガンドと非常に特異的に結合します。 化学的性質のために、糖タンパク質はより重要です。
受容体は機能を勝ち取ります:
1)タンパク質とイオンチャネルのコンフォメーションを変化させることにより、原形質膜の浸透を調節します。
2)クリチン中の特定の分子の供給を調節します。
3)センサーのように動作し、信号を内部信号に変換します。
4)ポサクリチンマトリックスの分子を細胞骨格に結合させる。 気受容体は呼ばれます インテグリン、悪臭は、クリチンとクリチンの間の接触の形成と間質性発話を確実にします。
原形質膜受容体は5つのファミリーに分けることができます:
- チャネルに関連する受容体 、vzaimodiyutのリガンド-神経伝達物質。timchasovovіdkrivaєまたはkomіrnymehanіzmを閉じ、その結果、チャネルを介したイオンの輸送がブロックされます。 チャネル形成受容体は、イオンを特異的に通過させる関連タンパク質サブユニットで構成されています。 グルタミン酸、γ-アミノ酪酸、グリシン、サイクリックモノヌクレオチド(cAMP、cGMP)はこれらの受容体と相互作用します。
- 触媒受容体 細胞内部分(信号を受信する水受容体)とプロテインキナーゼのような細胞質部分が含まれます。 シグナル伝達分子の形成により、クリチンの生化学的変化のカスケードが穂軸にもたらされ、それが歌う生理学的反応につながります。 インスリン、表皮および血小板成長因子、および神経成長因子がこれらの受容体に追加されます。
- Gタンパク質に結合した受容体 -酵素によってイオンチャネルに結合した膜貫通タンパク質。 分子のセシリウム複合体。これには次のものが含まれます。
1)シグナル分子(最初の中間体)と相互作用する受容体自体-原形質膜を7回貫通する中心タンパク質、これらの受容体の内部ループは、Gタンパク質の結合の中心を遮断します(たとえば、 β-アドレナリン受容体);
2)Gタンパク質(デカール成分で構成されるグアノシン三リン酸結合調節タンパク質)は、膜結合酵素(アデニル酸シクラーゼ)または イオンチャネル、その後アクティブ化されます。
3)別の内部細胞メディエーター-ほとんどの場合、サイクリックAMPまたはGMP(cAMP、gAMP)またはCa2+。
このような受容体を介して、神経伝達物質の80%、ペプチドホルモンの効果が実現されます。
-免疫グロブリン受容体-ce受容体-マクロファージおよび免疫担当細胞の表面にある免疫グロブリン。これにより、すべての外来生物および免疫生物の認識が保証されます。
-インテグリン-クリチン接着分子-サブクリチン線維性高分子の受容体として機能する膜貫通タンパク質-フィブロネクチンとラミニン。 フィブロネクチンは、クリチンおよびポストクリチンマトリックスの分子(コラーゲン、ヘパリン、フィブリン)に結合します。 細胞と細胞間発話の間の接着部位としてのフィブロネクチン。 インテグリンの細胞内部分は、他のタンパク質(ビンキュリン、タリン、α-アクチニン)を介して細胞骨格と接続しています。
このランクでは、原形質膜受容体は受け取ります 異なる信号必要に応じて、細胞の代謝を変化させ、細胞の速度、分泌を開始および調節し、膜の表面の電位を調節します。
細胞内受容体。細胞内受容体は、細胞の遺伝子活性を調節するタンパク質です。 悪臭は腐っています:
細胞質は膜に細胞小器官を持っています。 細胞質受容体は、ステロイドホルモン、例えば、グルコース、鉱質コルチコイド、アンドロゲン、およびプロゲステロンについて検出されます。 甲状腺ホルモンのミトコンドリア受容体;
核内-甲状腺ホルモンの核内受容体、エストロゲン、ビタミンD、レチノイン酸の受容体。
ステロイドホルモンの受容体には3つのドメイン(部分)があります:
1)ホルモン結合-リガンドとの相互作用のため;
2)DNA結合;
3)転写を活性化するドメイン。
このような疎水性受容体のシグナル分子は、原形質膜を通って自由に拡散し、次に内部の細胞タンパク質受容体に結合します。 この後、タンパク質のコンフォメーションが変化し、その活性化が起こり、胞子形成がDNAに移動します。 このようなホルモン受容体複合体は、核内の特定の遺伝子と結合し、それらの発現を調節して、細胞の機能状態を変化させる低酵素の生合成を確実にします。
テーマ5
MIJKLITOCHNI SOLUKI、タイプI構造-
機能分類
細胞間空間は特別な構造であり、原形質膜とともに、クリチン間の相互作用を提供します。 細胞間接触は糖衣によって保護され、タンパク質によって結合されます。 中間セクションは、2つの主要なタイプに細分できます。
1.Mehanichnіspoluky-クリチンの機械的リンクを1つずつ固定します。 それらの前に、単純で折りたたまれた折り目があります:schіlnіspoluki(schіlnycontact)、desmosomes、interdigitation。
2.コミュニケーションspoluky-クライアント間の安全な化学的コミュニケーション。 それらまで、あなたはschіlinnіspolkiを見ることができます。
メカニカルスラブ
I.単純な性交-15〜20nmの距離でのsucidclitinのプラズモレムの近似。 susidnіhクリチンのこの糖タンパク質を使用すると、特定の細胞が互いに「認識」し、受容体(カドヘリン、インテグリン)をトブトします。 Obov'yazkovoyumovoyєイオンCa2+の存在。 たとえば、E-cadheriniは、接触しているすべての表面に上皮細胞を確実に形成します(図3)。
図3-単純な断面接続(スキーム):