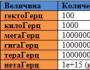
膜の受容体機能は タンパク質の受容体機能。 ホルモンシグナルの細胞内伝達の主なシステム
細胞の受容体機能は、同じように患者の反応を実現するために使用される受容体によって保証されます。
転送された情報から包帯を注入する方法。これは、スピーチの出現によるものであり、細胞の中央にある膜の受容体複合体からの呼び出しがあるはずです。
イオノトロピック受容体複合体は、イオンチャネルに加えて、折りたたまれた分子および超分子サブレイヤーを形成します。 生物学的に活発なスピーチに関連する場合、それは次のように聞こえます イオンチャネル。 Shvidkіstはklitini寺院を損傷しました。 イオノトロピック受容体はシナプスの領域で最も重要であり、興奮性および電気性滲出液の伝達に関与します。
代謝型受容体複合体は、不可欠な中間タンパク質と結合し、情報を内面に伝達します。 NasamperedtseGタンパク質および膜チロシンキナーゼ。 中間タンパク質はクリチン膜の内面の酵素を刺激し、tiはそれ自体で他の中間体を合成します-低分子量のスピーチはクリチンの生物学的反応を引き起こします。 気受容体は時々最も重要と呼ばれます。 同様のメカニズムを通じて、より多くのホルモンとメディエーターが、ひどく同様に、クリチンに浸透します。
低脂肪リポタンパク質倉庫の脂質など、細胞内の分子の供給を調節する受容体。 このグループの受容体は浸透を変えることができます 生体膜、そのようなランクで唾を吐く 化学倉庫クリチンの真ん中で。
接着性受容体(インテグリン、カドヘリン、免疫グロブリン、セレクチンなどの発祥の地)は、感受性クリチンまたはクリチンを細胞間媒体の構造、例えば基底膜と結合します。 接着相互作用の可能性は、細胞と生物全体の生活において重要である可能性があります。 接着に対するクリチニティの喪失は、制御されていない移動(転移)および分化障害を伴う。 タマン悪性肉付き細胞における接着受容体の機能の病理学的障害。
受信の上位プロセスは、特別な糖タンパク質である受容体の助けに依存しています。 悪臭は膜上球-糖衣細胞を歩き回っています。
受容体は、ホルモン、生物学的に活性な発話、感受性細胞の膜、細胞間発話の接着分子などの特定の細分化の付着に関与しています。 受容体は、クリチンの高度に特殊化された構造です。 悪臭は、特異性が高い(親和性が高い)場合もあれば、特異性が低い(親和性が低い)場合もあります。 特異性の程度は、クライアントの感度のレベルを決定します。 ホルモンの最も親和性の高い受容体。
受容体複合体はまた、膜の内部球の特徴である。 悪臭は、膜および非膜オルガネラ、カリオールの内側骨盤葉および皮膚に見られます。
特定の信号(スピーチレギュレーターを備えた受容体の受信)に応答して、それは生化学的反応の槍によるものであり、それは生物学的応答の形成につながります-細胞の目覚めまたは亜鉛メッキ。 ポリペプチド、類似のアミノ酸、抗原複合体、糖タンパク質などの受容体。 クリチンの膜にあります。 デヤキ受容体は、他のメディエーターの確立を確実にするタンパク質、およびイオンチャネルのタンパク質と結合することができます。 このような受容体システムは代謝型と呼ばれます。
代謝型受容体への損傷、viklikana信号は、細胞の内部にデキルコマの方法で伝達される可能性があります。 ある方法では、受容体とシグナル分子との相互作用により、受容体の立体配置が変化し、いわゆるGタンパク質の構造が変化し、それ自体が、シグナル分子の除去を活性化します。細胞質(他の中間体)。
Єアデニル酸シクラーゼとcAMPを活性化するGsタンパク質、アデニル酸シクラーゼを阻害するGiタンパク質、ホスホリパーゼCを活性化して細胞質ゾルのカルシウムイオンを増加させるGpタンパク質。 サイクリックグアノシン一リン酸(cGMP)のホスホジエステラーゼを活性化し、cGMPの存在を減少させ、細胞の亜鉛メッキ(膜過分極)を引き起こすGtタンパク質も知られています。 サイクリックAMP(cAMP)は、プロテインキナーゼを活性化し、クリチンの生化学反応を促進します。
別の例では、受容体は、Ras-Gタンパク質を活性化し、Rasカスケードをトリガーするチロシンキナーゼと結合します。 このプロセスの結果として、イノシトール-1,4,5-三リン酸、ジアシルグリセロールが溶解します。 Tseviklikaєlantsyug触媒反応、zokrema転写。
受容体は、イオンチャネルと関連し、それらの浸透を変化させ、膜の脱分極を振動させ、カルシウムイオンのクリチナへの浸透などを行うことができます。 悪臭は白いエフェクターの別棟、つまりイオンチャネルにやってくる。 不活性化酵素は、他のシグナルスピーチのメディエーターのシグナル分子から受容体結合を開きます。
受容体の一部のシグナル伝達機能のいくつかは、接着および凝集において重要な役割を果たします-細胞構造および/または細胞間構造と同様の、それ自体へのクリチンの接着。 胞子細胞の糖衣の受容体による「摂取」は、1時間の凝集を伴う。 このような受容体が個人、臓器、組織の特異性を示すことが重要です。 お尻はbutiselectini、integrini、cadheriniにすることができます。 それらはクリチンに抗原力を与え、それらが一つずつ「知る」ことを可能にします。
許しを知っているなら、親切にして、テキストの断片を見て、それを押してください Ctrl +Enter.
Be-yakіkіtinimаyut膜タンパク質-受容体。 これらは、脂質二重層を1回以上変換する膜貫通型タンパク質です。 それらにはオリゴ糖が含まれます(このような受容体はより正確には糖タンパク質と呼ばれます)。 このようなタンパク質は、外膜と豊富な細胞内膜の両方に存在します。 たとえば、小胞体の膜上のリアノジン受容体とイノシトール三リン酸受容体。 膜受容体はシグナルスピーチ(リガンド)に結合し、それらのコンフォメーションを変化させます。 イオンチャネルを使用した1時間の一部。 リガンドを結合するとき、チャネルはカールまたはカールする可能性があります。 このようなタンパク質は、イオノトロピック受容体と呼ばれます。 他の受容体は、リガンドに結合すると、膜の内側で化学反応を引き起こします(したがって、酵素または調節タンパク質と同時に悪臭を放ちます)。 このようなタンパク質は代謝型受容体と呼ばれます。 代謝型受容体の2つの主要なタイプ-Gタンパク質に結合したze受容体とプロテインキナーゼ活性を持つ受容体。 Gタンパク質に結合する受容体はセミヘリックス(アルファヘリックスの出現時に同時に膜を折りたたむ)タンパク質です。 ベータ2アドレナリン受容体の適用における作用機序を見てみましょう。 Tse-アドレナリンに敏感なアドレナリン受容体のタイプの1つ(ノルエピネフリンはそれらにとってそれほど重要ではありません)。 ピースリガンドの1つに結合したβ2-アドレナリン受容体の構造は、追加のX線回折分析によって明らかになりました。 気にアドレナリンが付着すると、気管支の滑らかな粘膜の受容体と骨格膜の血液を運ぶ血管が弱まり、肝臓の細胞ではグリコーゲンの分解(グリコーゲン分解)が増加し、グルコースが承認され、血液から出ます。 Gsタンパク質に結合するZimタイプの受容体。 このタンパク質は、他のGタンパク質と同様に、3つのサブユニット(ポリペプチドランス)(α、β、γ)で構成されています。 酪酸の2つの尾と膜表面の自由なperezovaetsyaの助けを借りて膜の内側に「追加」します。 不活性なGsタンパク質のαサブユニットはGDP分子に結合します。 ベータ2受容体がアドレナリンの影響を受けると、受容体が活性化され(そのコンフォメーションが変化し)、Gsタンパク質が活性化されます。 その結果、αサブユニットはβγサブユニットとして加水分解し、GDP分子をGTP分子と交換します。 このような活性なαサブユニットは、膜貫通型タンパク質である酵素アデニル酸シクラーゼと結合し、ヨガを活性化します。
Gsタンパク質とアデニル酸シクラーゼの活性化
この酵素は、ATPからのサイクリックアデノシン一リン酸(cAMP)の合成を仲介します。 CAMFは、細胞内の信号伝送に使用されるユニバーサルな2番目の仲介者の1つです。 で この特定のタイプに cAMPは、プロテインキナーゼの1つであるプロテインキナーゼA(PKA)を活性化します。 この酵素は4つのサブユニットで構成されています-2つは調節性で2つは触媒性です。 サイクリックAMPのchotyrox分子を結合するとき、調節サブユニットは触媒サブユニットとして水クリーム状になり、それらが活性化されるとき。 肝細胞では、RKAは別のプロテインキナーゼであるホスホリラーゼキナーゼをリン酸化します。 ホスホリラーゼキナーゼ、それ自身の黒色ホスホリラーゼ、グリコーゲンホスホリラーゼ。 ホスホリラーゼの作用下で、グリコーゲンのリン酸分解が起こります。 その結果、ブドウ糖はutvoryutsyaであり、タンパク質を介してヤクになります-キャリアは血液中の肝臓のクリチンから出て、spozhivaetsya器官はストレスに積極的に働きます-そもそも骨格のm'yazami。
プロテインキナーゼA(PKA)の活性化と不活性化のメカニズム。 PDE-ホスホジエステラーゼ。
そのような折り畳み可能でアクセスしやすい信号伝送システムの必要性は何ですか? まず、最初の段階で、信号はから送信されます 外側内側の膜(ヤクジホルモン用)、cAMPの合成に専念。 高分化型の親水性cAMP分子は、細胞全体に急速に拡散し、すべての細胞にシグナルを伝達します。 別の方法では、シグナル伝達の皮膚段階を調節することができます。 エール、ステージが大きいほど強い信号が必要になるため、頻繁に送信することの重要性は他の人にとっても重要です。 したがって、1時間の受容体活性の下で、Gタンパク質の非人格的な分子を活性化することができます。 皮膚のGsタンパク質は1分子のアデニル酸シクラーゼを活性化し、エールのアデニル酸シクラーゼは数千のcAMP分子を合成します。 4分子のcAMPは、PKAの2つの触媒サブユニットすべてを活性化しますが、他の分子は非ホスホリラーゼキナーゼ分子をリン酸化することができます。 このようなバガトラーゼ強化システムの結果として、1つのアドレナリン分子の下で、約10,000,000個のグルコース分子が肝細胞に吸収されます。 ストレスが過ぎると、アドレナリンの分泌が減少します。 アドレナリンは血液からニクを介して容易に排泄され、受容体への作用を停止します。 その後、Gsタンパク質は不活化されます。 Gタンパク質はGTP-ase活性を変化させます:α-サブユニットはGTP分子をGDP(iリン酸)に分解し、その後βγ-サブユニットに結合して不活性状態に移行します。 そのようなランクでは、Gタンパク質は自動の「分子ビミック」のようなものです。 蛹のcAMPのルバーブは、cAMPが特定の酵素であるホスホジエステラーゼを分解するという事実の手がかりであると考えられています。 その結果、RKAの触媒サブユニットはレギュレーターと結合して不活性化されます。 キナーゼホスホリラーゼおよびホスホリラーゼグリコーゲンの不活性化は、プロテインホスファターゼ酵素によって阻害されます。プロテインホスファターゼ酵素は、これらの酵素をリン酸基に分割します。 そのため、グリコーゲンの分解が付着しています。
 生物の基礎の心の1つは、クリチン内のすべての化学的および物理的プロセスのオーバーフローの使用です。 進化の過程で、これらの規制メカニズムの組み合わせがあり、生物学的システムにおける物理的および化学的プロセスの最も効率的な使用を保証しました。 豊富な細胞生物では、生物のバランス、組織活動の調整、および進化の過程でのクリチンの生理学的機能の制御された調節が、細胞間相互作用の折り畳みメカニズムを振動させ、分子の用量を実現するのに役立ちました。
生物の基礎の心の1つは、クリチン内のすべての化学的および物理的プロセスのオーバーフローの使用です。 進化の過程で、これらの規制メカニズムの組み合わせがあり、生物学的システムにおける物理的および化学的プロセスの最も効率的な使用を保証しました。 豊富な細胞生物では、生物のバランス、組織活動の調整、および進化の過程でのクリチンの生理学的機能の制御された調節が、細胞間相互作用の折り畳みメカニズムを振動させ、分子の用量を実現するのに役立ちました。
 細胞の機能を調節する方法:1。酵素の活性の変化2.発話の区画化の変化。 3.地形力学的調節。 4.タンパク質の化学修飾。 5.ゲノムの発現。 6.二次仲介者の活性化。
細胞の機能を調節する方法:1。酵素の活性の変化2.発話の区画化の変化。 3.地形力学的調節。 4.タンパク質の化学修飾。 5.ゲノムの発現。 6.二次仲介者の活性化。
 1.酵素の活動の変化の心の細胞機能の調節の最初の方法。 2.真核生物における代謝過程の区画化(空間床)の役割はすでに大きい。 サイトゾルおよび他の細胞小器官における特定の代謝プロセスの局在化は、これらのプロセスの独立した調節を容易にし、代謝のより微妙な調節を可能にします。 発酵の破片と基質はしばしば膜によって分離され、基質の透過性を変化させることは、間違いなく、1時間の酵素反応を意味します。
1.酵素の活動の変化の心の細胞機能の調節の最初の方法。 2.真核生物における代謝過程の区画化(空間床)の役割はすでに大きい。 サイトゾルおよび他の細胞小器官における特定の代謝プロセスの局在化は、これらのプロセスの独立した調節を容易にし、代謝のより微妙な調節を可能にします。 発酵の破片と基質はしばしば膜によって分離され、基質の透過性を変化させることは、間違いなく、1時間の酵素反応を意味します。
 3. У зв'язку з тим, що багато ферментів клітини вбудовані в цитоплазматичні мембрани, існує третій шлях регуляції клітинної активності – так звана топодинамічна регуляція, що ґрунтується на динамічній неоднорідності розподілу білків у мембрані, їх здатності утворювати асоціації. 高分子複合体における代謝反応の変異配列を触媒する酵素の会合は、それらの仕事を調整し、代謝経路における中間体の安全な動きを確実にすることを可能にします。
3. У зв'язку з тим, що багато ферментів клітини вбудовані в цитоплазматичні мембрани, існує третій шлях регуляції клітинної активності – так звана топодинамічна регуляція, що ґрунтується на динамічній неоднорідності розподілу білків у мембрані, їх здатності утворювати асоціації. 高分子複合体における代謝反応の変異配列を触媒する酵素の会合は、それらの仕事を調整し、代謝経路における中間体の安全な動きを確実にすることを可能にします。
 4.細胞活性を調節する4番目の方法は、酵素や他のタンパク質の化学修飾です。 化学基がタンパク質に来るという事実に応じて、それらはアセチルウバニヤ(過剰のオクト酸CH 3 CO-を導入する)、アデニルバニヤ、水酸化物、メチル化、ホスホリルバニヤなどを区別します。 化学基の追加は、タンパク質分解に対する耐性など、タンパク質の機能的活性と力を変化させます。 タンパク質(ヒドロキシルバニヤ、メチルユバニヤ)のデヤキ修飾は、長い寿命によって処理され、そうでなければ、長い間、デブ(アセチルウバニヤ、アデニルバニヤ)、フビリン(リン酸化)によって処理されます。 タンパク質のリン酸化を達成するための最良の方法は、リン酸基のタンパク質に追加することです。
4.細胞活性を調節する4番目の方法は、酵素や他のタンパク質の化学修飾です。 化学基がタンパク質に来るという事実に応じて、それらはアセチルウバニヤ(過剰のオクト酸CH 3 CO-を導入する)、アデニルバニヤ、水酸化物、メチル化、ホスホリルバニヤなどを区別します。 化学基の追加は、タンパク質分解に対する耐性など、タンパク質の機能的活性と力を変化させます。 タンパク質(ヒドロキシルバニヤ、メチルユバニヤ)のデヤキ修飾は、長い寿命によって処理され、そうでなければ、長い間、デブ(アセチルウバニヤ、アデニルバニヤ)、フビリン(リン酸化)によって処理されます。 タンパク質のリン酸化を達成するための最良の方法は、リン酸基のタンパク質に追加することです。
 一般にタンパク質分子にリン酸基を付加する酵素は、プロテインキナーゼと呼ばれます。 Dzherelomリン酸基єATP。 逆反応(脱リン酸化)はプロテインホスファターゼの影響を受けます。 5.細胞活動を調節する5番目の方法として、ゲノムの発現を呼び出す必要があります。 気候におけるタンパク質の量の調節は、その合成の誘導および抑制によって、すなわち、遺伝子の転写速度の変化の結果として影響を受ける。 豊富な細胞の真核生物では、誘導物質の役割は、甲状腺ホルモンやステロイドホルモンなどの特定の調節因子によって果たされます。
一般にタンパク質分子にリン酸基を付加する酵素は、プロテインキナーゼと呼ばれます。 Dzherelomリン酸基єATP。 逆反応(脱リン酸化)はプロテインホスファターゼの影響を受けます。 5.細胞活動を調節する5番目の方法として、ゲノムの発現を呼び出す必要があります。 気候におけるタンパク質の量の調節は、その合成の誘導および抑制によって、すなわち、遺伝子の転写速度の変化の結果として影響を受ける。 豊富な細胞の真核生物では、誘導物質の役割は、甲状腺ホルモンやステロイドホルモンなどの特定の調節因子によって果たされます。
 6.クリチンシステムの活動を調節する最良の方法では、それはいわゆる第二の仲介者(仲介者、メッセンジャー)と接続されています。 その結果、クリチンの中央にあるシグナル分子と膜受容体の間の相互作用が二次中間体のランクを振動させ、カスケードメカニズムを引き起こし、クリチンの活性に変化をもたらします。
6.クリチンシステムの活動を調節する最良の方法では、それはいわゆる第二の仲介者(仲介者、メッセンジャー)と接続されています。 その結果、クリチンの中央にあるシグナル分子と膜受容体の間の相互作用が二次中間体のランクを振動させ、カスケードメカニズムを引き起こし、クリチンの活性に変化をもたらします。
 シグナル伝達分子を参照してください:膜を横切って拡散し、細胞内受容体に結合する小さな親油性分子。 細胞膜受容体と相互作用する親油性分子。 細胞膜受容体(親水性ホルモン、成長因子、サイトカイン、神経伝達物質)と相互作用する親水性分子。
シグナル伝達分子を参照してください:膜を横切って拡散し、細胞内受容体に結合する小さな親油性分子。 細胞膜受容体と相互作用する親油性分子。 細胞膜受容体(親水性ホルモン、成長因子、サイトカイン、神経伝達物質)と相互作用する親水性分子。
 最初のグループには、関節のひだと皮膚のひだの樹皮によって振動するステロイドホルモンが含まれています。 ステロイドホルモン受容体は、リガンドに対して非常に振動している可能性があります。 悪臭は、ホルモンを含まない大きなタンパク質(50〜120 k)で表され、細胞質または核に広がり、多くのドメイン、ヤクvykonuyutの歌う機能を含みます。
最初のグループには、関節のひだと皮膚のひだの樹皮によって振動するステロイドホルモンが含まれています。 ステロイドホルモン受容体は、リガンドに対して非常に振動している可能性があります。 悪臭は、ホルモンを含まない大きなタンパク質(50〜120 k)で表され、細胞質または核に広がり、多くのドメイン、ヤクvykonuyutの歌う機能を含みます。
 シグナル伝達分子の他のグループには、アラキドン酸の誘導体であるエイコサノイド(ギリシャ語のエイコシ-20からの酸化された多価不飽和脂肪酸)が含まれます。 それらの代表は、プロスタグランジン、ロイコトリエン、プロスタサイクリンおよびトロンボキサンです。 エイコサノイドはすべてのクリチンで実際に承認されており、その生合成はホスホリパーゼA2によって開始されます。
シグナル伝達分子の他のグループには、アラキドン酸の誘導体であるエイコサノイド(ギリシャ語のエイコシ-20からの酸化された多価不飽和脂肪酸)が含まれます。 それらの代表は、プロスタグランジン、ロイコトリエン、プロスタサイクリンおよびトロンボキサンです。 エイコサノイドはすべてのクリチンで実際に承認されており、その生合成はホスホリパーゼA2によって開始されます。
 シグナル伝達分子の3番目のグループは、親水性ホルモン、成長因子、リンホカイン(サイトカイン)で構成されています。 ホルモンは、特別な内分泌沈着物またはクリチンによってviroblyayutsyaであり、追加の血流のためにクリチン標的に輸送されます。 成長因子とサイトカインの両方が組織ホルモンのグループと結合します。 Spіlnimїmєtak:viroblyayutsyaかなりの非内分泌細胞。 隙間空間での拡散経路によって拡大します。 マユットパラクリンまたはオートクリン地獄。
シグナル伝達分子の3番目のグループは、親水性ホルモン、成長因子、リンホカイン(サイトカイン)で構成されています。 ホルモンは、特別な内分泌沈着物またはクリチンによってviroblyayutsyaであり、追加の血流のためにクリチン標的に輸送されます。 成長因子とサイトカインの両方が組織ホルモンのグループと結合します。 Spіlnimїmєtak:viroblyayutsyaかなりの非内分泌細胞。 隙間空間での拡散経路によって拡大します。 マユットパラクリンまたはオートクリン地獄。
 成長因子はタンパク質であり、ポジール(ingibuyut)と歌う細胞の発達を刺激します。 成長因子表皮成長因子を適用します。 ニューロン成長因子; 線維芽細胞(細胞後マトリックスを合成する体の健康な組織内の細胞)の成長の要因。 サイトカインは、体の発火、免疫およびその他の免疫反応に関与します。 シムに関連して、悪臭は免疫系のクリチンによって振動されます。 インターロイキンとインターフェロンは、サイトカインのお尻として機能することができます。
成長因子はタンパク質であり、ポジール(ingibuyut)と歌う細胞の発達を刺激します。 成長因子表皮成長因子を適用します。 ニューロン成長因子; 線維芽細胞(細胞後マトリックスを合成する体の健康な組織内の細胞)の成長の要因。 サイトカインは、体の発火、免疫およびその他の免疫反応に関与します。 シムに関連して、悪臭は免疫系のクリチンによって振動されます。 インターロイキンとインターフェロンは、サイトカインのお尻として機能することができます。
 二次仲介者によって条件付けられた細胞活動の調節のためのシステム。 異なるシグナル分子のシグナル伝達には4つのタイプがあります。
二次仲介者によって条件付けられた細胞活動の調節のためのシステム。 異なるシグナル分子のシグナル伝達には4つのタイプがあります。
 パラクリン調節の間、分泌細胞によって見られるシグナル伝達分子は、隣接する標的細胞に見られます。 シナプス伝達、神経系のみに特徴的で、誰にも見えません。 オートクリンシグナル伝達では、このクリチナによって生成されるシグナル伝達分子がそのクリチナに注入されます。 成長の豊かな要因を開発するためのTsimの方法。
パラクリン調節の間、分泌細胞によって見られるシグナル伝達分子は、隣接する標的細胞に見られます。 シナプス伝達、神経系のみに特徴的で、誰にも見えません。 オートクリンシグナル伝達では、このクリチナによって生成されるシグナル伝達分子がそのクリチナに注入されます。 成長の豊かな要因を開発するためのTsimの方法。
 追加のポストクリチンシグナル伝達分子の細胞間コミュニケーションには、いくつかの主要なステップが含まれます。1.シグナル伝達分子の生合成2.シグナル伝達分子の分泌3.シグナル伝達分子のクリチン標的への輸送4.特定の受容体によるシグナルの受信。 5.リガンド*-受容体複合体によって引き起こされる遺伝子の細胞代謝または発現の変化 *-リガンドの下では、シグナル分子がここで理解されます。
追加のポストクリチンシグナル伝達分子の細胞間コミュニケーションには、いくつかの主要なステップが含まれます。1.シグナル伝達分子の生合成2.シグナル伝達分子の分泌3.シグナル伝達分子のクリチン標的への輸送4.特定の受容体によるシグナルの受信。 5.リガンド*-受容体複合体によって引き起こされる遺伝子の細胞代謝または発現の変化 *-リガンドの下では、シグナル分子がここで理解されます。

 Klitinniy受容体-細胞質に豊富に存在するKlitinnyオルガネラであるKlitiniの表面にある分子で、歌われた分子の構成のヘビに特異的に反応し、膜貫通型イオンストリームなどの二次中間体を助けます。
Klitinniy受容体-細胞質に豊富に存在するKlitinnyオルガネラであるKlitiniの表面にある分子で、歌われた分子の構成のヘビに特異的に反応し、膜貫通型イオンストリームなどの二次中間体を助けます。
 受容体の特徴:1。選択性2.不在3.リガンドへのスポリティ。 4.組織特異性5.組織の可逆性
受容体の特徴:1。選択性2.不在3.リガンドへのスポリティ。 4.組織特異性5.組織の可逆性
 1.選択性。 Tsyaの力は、suvoriyの構造的特異性に基づいています。 このリガンドは、受容体に結合する単一の発話に関与するか、結合の場所を求めて他の人と効果的に競合します。 2.豊かさ。 zv'yazuvannyaの月の数はbutikіtsevim3である可能性があります。リガンドまでのSporidnіst。 違いは、アゴニストの生理学的濃度に依存します。 受容体を活性化する薬剤はアゴニストと呼ばれます。 拮抗薬を翻訳するための不活性なスタン受容体を持っている。
1.選択性。 Tsyaの力は、suvoriyの構造的特異性に基づいています。 このリガンドは、受容体に結合する単一の発話に関与するか、結合の場所を求めて他の人と効果的に競合します。 2.豊かさ。 zv'yazuvannyaの月の数はbutikіtsevim3である可能性があります。リガンドまでのSporidnіst。 違いは、アゴニストの生理学的濃度に依存します。 受容体を活性化する薬剤はアゴニストと呼ばれます。 拮抗薬を翻訳するための不活性なスタン受容体を持っている。
 4.組織特異性アゴニストの結合はこの組織で観察され、その生物学的効果に関与しています。 5.狼男dії。 アゴニストの受容体への結合の遮断は、可逆的な生物学的効果をもたらす可能性があります。
4.組織特異性アゴニストの結合はこの組織で観察され、その生物学的効果に関与しています。 5.狼男dії。 アゴニストの受容体への結合の遮断は、可逆的な生物学的効果をもたらす可能性があります。
 分類は、急性期後の信号の伝達メカニズムに基づいています。 アプローチに応じて、4種類の受容体が見られます。1。受容体チャネル。 リガンドと結合して受容体のコンフォメーションを変化させ、最初のイオンがチャネルを通過できるようにします。 そのような受容体の活性化の結果は、第2のメディエーターの細胞内レベルの膜電位の変化である。 適用:神経筋側のイオンチャネルとニコチン性アセチルコリン受容体。 筋小胞体のCa2+チャネル。
分類は、急性期後の信号の伝達メカニズムに基づいています。 アプローチに応じて、4種類の受容体が見られます。1。受容体チャネル。 リガンドと結合して受容体のコンフォメーションを変化させ、最初のイオンがチャネルを通過できるようにします。 そのような受容体の活性化の結果は、第2のメディエーターの細胞内レベルの膜電位の変化である。 適用:神経筋側のイオンチャネルとニコチン性アセチルコリン受容体。 筋小胞体のCa2+チャネル。
 2.タンパク質に結合した受容体。 リガンドが受容体に結合した後、Gタンパク質が活性化され、二次中間体を振動させる酵素を刺激または阻害します。 適用:コリン作動性、アドレナリン作動性および他。
2.タンパク質に結合した受容体。 リガンドが受容体に結合した後、Gタンパク質が活性化され、二次中間体を振動させる酵素を刺激または阻害します。 適用:コリン作動性、アドレナリン作動性および他。
 3.酵素活性を示す触媒受容体。 このクラスの受容体には、スプラットサブタイプが含まれます。グアニル酸シクラーゼ活性を示す受容体。 ホスファターゼ活性を示す受容体; チロシンキナーゼ活性を示す受容体。 サブグループの残りの部分には、成長因子が豊富なインスリン受容体があります。
3.酵素活性を示す触媒受容体。 このクラスの受容体には、スプラットサブタイプが含まれます。グアニル酸シクラーゼ活性を示す受容体。 ホスファターゼ活性を示す受容体; チロシンキナーゼ活性を示す受容体。 サブグループの残りの部分には、成長因子が豊富なインスリン受容体があります。
 4.触媒活性を示さない受容体は、チロシンキナーゼと関連しています。 リガンドとの結合は、単量体受容体の二量体化をもたらし、したがって、チロシンキナーゼを活性化する。 適用:サイトカイン、インターフェロン、成長因子の受容体。
4.触媒活性を示さない受容体は、チロシンキナーゼと関連しています。 リガンドとの結合は、単量体受容体の二量体化をもたらし、したがって、チロシンキナーゼを活性化する。 適用:サイトカイン、インターフェロン、成長因子の受容体。
 細胞受容体の別の分類:1。機械受容器(後で見える)2。熱受容器(視覚的に見えない)3。化学受容器a)イオノトロピックb)代謝型
細胞受容体の別の分類:1。機械受容器(後で見える)2。熱受容器(視覚的に見えない)3。化学受容器a)イオノトロピックb)代謝型
 化学受容器:1。受容体チャネル(図b); 2. Gタンパク質に結合する受容体(図a)。 3.酵素活性を示す受容体(図gzh)。 4.チロシンキナーゼ結合受容体(酵素活性は示しませんが)(図c)
化学受容器:1。受容体チャネル(図b); 2. Gタンパク質に結合する受容体(図a)。 3.酵素活性を示す受容体(図gzh)。 4.チロシンキナーゼ結合受容体(酵素活性は示しませんが)(図c)
 1.ブドバと受容体チャネルの分岐のメカニズム。 受容体-神経筋層のシナプス後膜にあるニコチン性アセチルコリン受容体のチャネル。 ニコチン性アセチルコリン受容体は糖タンパク質であり、4つの異なる遺伝子によってコードされる5つのサブユニット(2、)で構成されています。 アセチルコリンをリンクするための2本のボトルがあります。 アセチルコリンの2分子が受容体に結合すると、コンフォメーション変化が起こり、チャネルが開きます。 チャネルを開く時間は約1ミリ秒になります。
1.ブドバと受容体チャネルの分岐のメカニズム。 受容体-神経筋層のシナプス後膜にあるニコチン性アセチルコリン受容体のチャネル。 ニコチン性アセチルコリン受容体は糖タンパク質であり、4つの異なる遺伝子によってコードされる5つのサブユニット(2、)で構成されています。 アセチルコリンをリンクするための2本のボトルがあります。 アセチルコリンの2分子が受容体に結合すると、コンフォメーション変化が起こり、チャネルが開きます。 チャネルを開く時間は約1ミリ秒になります。

人体の機能は19世紀の初めを意識するようになりました。 Vcheniは、protosという単語の後に、ギリシャ語の「タンパク質」でスピーチの数を指定しました–「頭、最初」。
これらの化学組成の主な特徴は、新しい細胞が作られる前に代用する生物のように、基礎となるものです。 その他の機能は、安全な規制および交換プロセスに関連しています。 ビコナンニ輸送機能(例えば、血流から生物全体に広がるヘモグロビンタンパク質); m'yazovih繊維の成形; 体の重要な機能の管理において(私たちはインスリンタンパク質のお尻を使用します); エッチングプロセスの調整、エネルギー交換; 体の防衛で。
これらのスピーチの化学構造は、タンパク質分子の数を含むアミノ酸の数によって決定されます。 世界を超えた分子は大きいです。 ペプチド結合によって結合されたスピーチの数、高分子有機スピーチ、およびアミノ酸のランス。 遺伝暗号に関連するタンパク質のアミノ酸貯蔵。 アミノ酸の数の豊富なバリエーションは、タンパク質分子のさまざまな力を与えます。 原則として、悪臭は互いに混ざり合い、複雑な複合体を作ります。
タンパク質の分類は十分に活用されておらず、すべてのタンパク質から遠く離れて科学者によって研究されてきました。 それらの多くの役割は、人々にとって謎のままです。 当分の間、タンパク質はそれらの生物学的役割とアミノ酸自体がそれらの倉庫に入るという事実に従って分けられます。 私たちの食事にとって、亜鉛はタンパク質そのものではなく、ヨガのアミノ酸です。 アミノ酸はさまざまな有機酸の1つです。 それらは100以上あり、それらがなければ代謝プロセスを実行することは不可能です。
体はそれに付随するタンパク質を取り戻すことはできません。 それらのほとんどは、酸っぱいハーブジュースの影響下にあります。 タンパク質のアミノ酸への分解が観察されます。 生物は、必要なアミノ酸の分解後に「取り込み」、必要なタンパク質を構築します。 この場合、一部のアミノ酸から他のアミノ酸への変換が期待できます。 変容のクリーム、悪臭は生物で独立して合成することができます。
すべてのアミノ酸の通過は私たちの体を振動させることができます。 Ti、yakіは合成されていません、それらはかけがえのないものと呼ばれているので、体はそれらを必要とします、しかしそれらは呼び出すことができるだけです。 必須アミノ酸を他のアミノ酸に置き換えることはできません。 それらには、メチオニン、リジン、イソロイシン、ロイシン、フェニルアラニン、スレオニン、バリンが含まれます。 それまでは、他のアミノ酸は、必須のフェニルアラニンとメチオニンのみからのutvoryuyuyutsyaです。 このため、食事の質は、入手可能な白の量ではなく、倉庫によって決まります。 たとえば、ジャガイモ、白キャベツ、ビートルート、キャベツ、マメ科植物、パンには、大量のトリプトファン、リジン、メチオニンが含まれています。
私たちの生物へのタンパク質交換の通過は、必要なタンパク質の十分な量で堆積されるべきです。 一部のスピーチの分割と他のスピーチへの変換は、生物のエネルギー消費に依存します。
体の活力の遺産として、白人の一部の無駄が絶えず観察されています。 呼び出す必要のあるタンパク質のスピーチから、約30gが生産に費やされます。 そのために、改善にお金を費やし、食事療法は、体の練習を確保するために、これらのスピーチの十分な復讐の罪を犯しています。
さまざまな要因のために横になる白いスピーチの本体によるSpozhivannya:vikonnannya重要な身体的仕事または平和の駅での休息。 感情的な状態。 獲物の場合、タンパク質の保持率は成人の場合50グラム未満である必要があります(体重1キログラムの皮膚あたり約0.8グラム)。 成長率が高い子供には、より多くのタンパク質が必要です-体重1キログラムあたり最大1.9グラム。
ティムも少なくありません。ハリネズミに慣れているタンパク質のスピーチがたくさんあり、それらのアミノ酸のバランスの取れた数を保証するものではありません。 したがって、食事の食事は異なる可能性があり、その結果、体はさまざまなアミノ酸から最大限の腐食性を得ることができます。 今日でもトリプトファンを見せなかった人を見つけることはできません。明日は病気になります。 Ні、有機体は、消費のさまざまな時間に腐食性アミノ酸とガラス製品を保存するために少量で「インテリジェント」です。 ただし、体の累積的な健康状態はそれほど高くないため、茶色のスピーチのストックは定期的に補充する必要があります。
Yakshchoは、個人の場合、過剰な再エナミ(菜食主義)は、Healthyの村(同じうなり声の領域であるZi Shlunkovo-Kishkovykhの問題)を棄権します。 likar-sibliyt、彼の汚れたSkoriguvatiは、Protheanのバランスのバランスです。
集中的なスポーツ活動では、体は大量のタンパク質を必要とします。 そのような人々のために特別に、スポーツ食品が生産されています。 Protenadkhodzhennyaproteinіvは、vykonuyutsyaの物理的な衝動を示している可能性があります。 しかし、考えが広がったにもかかわらず、これらのスピーチが多すぎると、マゾボイの質量が急激に増加することはありませんでした。
タンパク質機能の多様性は、体内で発生するすべての生化学的プロセスに限定されません。 それらは生化学的触媒と呼ぶことができます。
タンパク質から、細胞骨格が確立され、クリチンの形態をサポートします。 タンパク質がなければ、免疫系の機能は不可能です。
Vіdmіnnykharchovimdzherelomタンパク質єm'yaso、ミルク、魚、シリアル、マメ科植物、エンドウ豆。 たんぱく質が少ない果物、ベリー、野菜。
最初のタンパク質は、そのアミノ酸配列を指定するための一種の文字であり、インスリンです。 この功績により、F。センジャーは前世紀の60年間でノーベル賞を受賞しました。 また、D。KendryuとM. Perutzの科学者は、X線回折の追加技術を使用して、ミオグロビンとヘモグロビンのトリビミック構造を同時に作成することができました。 その価格で彼らはノーベル賞も受賞しました。
イベントの歴史

タンパク質学会の創設者はアントワーヌ・フランソワ・ド・フルクロワです。 okremiyクラスのvidilivїхで、その後、ヤクpominіvіvіvіstіdenaturirovat(またはgortatisya)はvysokїї温度で酸を加えました。 Vin doslidzhuvavフィブリン(血液からの視覚)、グルテン(小麦粒からの視覚)およびアルブミン(卵白)。
オランダの学者G.Mulderは、フランス人の同僚であるde Fourcroixの科学的研究を完了し、タンパク質倉庫の分析を実施しました。 ワインのこの分析から、タンパク質分子の大部分が実験式に似ている可能性があるという仮説を立てました。 Vinは、タンパク質の分子量を最初に決定した人物でもあります。
Mulderによれば、どのタンパク質も小さな構造倉庫、つまり「タンパク質」で構成されています。 そして1838年、J。ベルセリウスは、すべてのタンパク質の一般名として「タンパク質」という用語を提唱することにより、スウェーデンの教えを推進しました。
30〜40歳の発症時に、タンパク質倉庫に入るアミノ酸の大部分の研究が行われました。 1894年、ドイツの生理学者であるA. Kosselは、アミノ酸自体と非常に構造的な貯蔵タンパク質、そして悪臭がペプチド結合によって結合されていることを認めました。 タンパク質のnamagavsyadoslіdzhuvatiアミノ酸配列。
1926年に、roci、nareshti、および体内のタンパク質の主な役割が認識されました。 その後、Tseは、米国のD. Sumnerの化学者がそうした場合、そのウレアーゼ(酵素、それなしでは豊富な化学プロセスを克服することは不可能でした)єタンパク質になりました。
当時の科学の必要性のために純粋なタンパク質を見ることはさらに折り畳み可能でした。 同じ理由で、これらのポリペプチドの接種により、最初の研究が実施されました。 最小限の硝子体血液タンパク質、鶏肉タンパク質、さまざまな毒素、ハーブおよび代謝酵素など、かなりの量から精製します。これらは非常に薄くなった後に見られます。 たとえば、1950年代には、ビチャッハ膵臓リボヌクレアーゼを精製することが可能でした。 スピーチ自体が学者の実験対象になりました。
科学では、タンパク質の研究が新しいレベルで続けられました。 これは、プロテオミクスと呼ばれる生化学の本質です。 さて、zavdyakiproteomytsі、あなたは精製されたタンパク質を見ることができるだけでなく、並行して、異なるクリチンや組織に見ることができる匿名のタンパク質の修飾の1時間の変化を見ることができます。 さまざまな科学者が、アミノ酸の配列によってタンパク質の構造を理論的に分解することができます。 低温電子顕微鏡法は、大小のタンパク質複合体の検出を可能にします。
たんぱく質の力
タンパク質の数は、合計するアミノ酸の数、または分子量を示すダルトンで測定できます。 たとえば、酵母タンパク質は450アミノ酸で構成されており、分子量は53キロダルトンです。 チチンと呼ばれる可能性のある最大の既知の現代科学タンパク質は、38000を超えるアミノ酸で構成されており、分子量は3700キロダルトンに近い。過剰なリン酸と相互作用するもののバランスをとるために核酸と結合するタンパク質は、主要なタンパク質によって考慮されます。 プロタミンとヒストンはそれらの前に見ることができます。
リスは多様性の平等に応じて分けられ、その大部分は水によってよく区別されます。 traplyayutsyaとブドウの木を保護します。 フィブロイン(ウェブと継ぎ目の基礎)とケラチン(人間の髪の毛の基礎、および生き物や鳥の羊毛の基礎)は不明瞭です。
変性
原則として、タンパク質は、そのような悪臭に対して、生物の構造の物理的および化学的力を保持します。 その後、体が歌う温度にしがみつくと、タンパク質が見え、その優位性は変わりません。そのような心の変化は、余分な温度のようなものです、またはそれが真ん中の酸/水たまりで消費される場合、それはタンパク質が2番目、3番目および4分の1の構造を消費するという事実につながる可能性があります。 強力な生きているクリチンである本来の構造の喪失は、タンパク質摂取の変性と呼ばれます。 変性は、私的または再発性、非可逆性または可逆性である可能性があります。 交渉不可能な変性の最も人気があり、お尻の端のバットは、鶏卵のクールな準備です。 高温下では、卵白アルブミン、プロソリウムタンパク質は不透明で狭くなります。
場合によっては、変性が逆転し、アンモニウム塩の助けを借りてタンパク質のリターンミルを回すことができます。 タンパク質精製の方法として逆変性を行っています。
シンプルで折り畳み可能なタンパク質
ペプチドランスのクリーム、いくつかのタンパク質の貯蔵には、非アミノ酸構造単位が含まれています。 可視性の基準、またはタンパク質内の非アミノ酸フラグメントの存在に従って、それらはフォールディングタンパク質と単純タンパク質の2つのグループに分けられます。 単純なタンパク質はアミノ酸ランスのみで構成されています。 折りたたみ可能なタンパク質は、非タンパク質の性質を破壊する断片を一掃します。化学的性質について 折りたたみバッグ 5つのクラスを参照してください。
- 糖タンパク質。
- 色素タンパク質。
- リンタンパク質。
- 金属タンパク質。
- リポタンパク質。
色素タンパク質は、フラボタンパク質、クロロフィル、ヘモグロビンなどを含むフォールディングタンパク質の一般的な名前です。
リンタンパク質と呼ばれるタンパク質は、倉庫から過剰なリン酸を取り除きます。 タンパク質グループのレベルには、たとえば、ミルクのカゼインがあります。
金属タンパク質-ceタンパク質、deiky金属のyakіmіstya共有結合イオン。 それらの中には、輸送および貯蔵機能(トランスフェリン、フェリチン)に関与するタンパク質があります。
リポタンパク質のフォールディングタンパク質は、過剰な脂質のために倉庫に保管する必要があります。 その機能は脂質の輸送です。
タンパク質の生合成
生物はアミノ酸に基づいてタンパク質を作ります 遺伝情報、ヤクは遺伝子にコード化されています。 タンパク質の合成による皮膚は、共役アミノ酸のユニークな配列で構成されています。 ユニークな配列は、タンパク質の販売に関する情報をコードする遺伝子のヌクレオチド配列などの要因によって決定されます。遺伝暗号はコドンで構成されています。 コドンは、過剰なヌクレオチドで構成される遺伝情報の単位です。 コドンからの皮膚は、タンパク質に1つのアミノ酸を追加するために組換えられます。 コドンの総数は64です。活性アミノ酸は1つではなく、いくつかのコドンによって割り当てられます。
体内のタンパク質の機能
他の多くの生物学的高分子(多糖類と脂質)は、クリチンのより大きな生命過程の発達のために体によって必要とされます。 タンパク質は代謝プロセスとエネルギー変換に影響を与えます。 悪臭は細胞小器官の倉庫に入ります-クリチン構造は、細胞間音声の合成に参加します。タンパク質の機能の分類は賢明であるため、同じタンパク質の一部の生物では少数の異なる機能を獲得できることを考慮に入れる必要があります。 タンパク質の豊富な機能は、高い酵素活性を持っているかもしれないもののベールを殺します。 Zocrem、そのような酵素には、ミオシンタンパク質、およびプロテインキナーゼ調節タンパク質が含まれます。
触媒機能
生物におけるタンパク質の最も重要な役割は、さまざまな化学反応の触媒作用です。 酵素は、特定の触媒力を持つタンパク質のグループです。 そのような酵素からの皮膚は、多くの同様の反応の1つの触媒です。 科学的には、何千もの酵素スピーチがあります。 たとえば、エッチングの過程でタンパク質を分解するスピーチペプシンは酵素です。私たちの体で起こる4,000以上の反応は触媒作用を必要とします。 酵素の流入がなければ、反応は数十倍、数百倍も進行します。
反応過程で酵素に付着し、その後修飾される分子は、基質と呼ばれます。 酵素にはアミノ酸の貯蔵庫がありますが、それらのすべてが基質と相互作用するどころか、さらに、それらのすべてが直接触媒プロセスに関与しているわけではありません。 基質が来る酵素のその部分は、活性酵素中心です。
構造機能
細胞骨格の構造タンパク質は、クリチンの形を与える一種のハードベースです。 Zavdyakiそれはクリチンの形を変えることができます。 それらの前に、エラスチン、コラーゲン、ケラチンを見ることができます。 成功した組織における細胞間発話の主な構成要素は、コラーゲンとエラスチンです。 ケラチンは、鳥だけでなく、髪や爪の基礎でもあります。Zahisna関数
Viriznyayutkіlkazahisnyhfunktsіyproteiniv:物理的、免疫的、化学的。物理的な成形では、コラーゲンが運命をたどります。 ワインは、骨、軟骨、腱、皮膚の深い球(真皮)など、さまざまな種類の成功した組織の細胞間発話の基礎を確立します。 血液に関与するタンパク質、トロンビン、フィブリノーゲンのグループのお尻。
免疫防御は、病原性微生物、またはposhkodzhennyaの攻撃に対する保護生物の形成において、血液プールまたは他の生物学的母親に入るタンパク質の運命を移します。 たとえば、免疫グロブリンはウイルス、細菌、または外来タンパク質を中和します。 免疫系によって振動する抗体は、抗原と呼ばれるため、体外の発話に付着し、中和します。 原則として、抗体は細胞間空間に分泌されるか、形質細胞の特殊な細胞の膜に固定されます。
酵素と基質は密接に関連していません。そうでないと、触媒反応が破壊される可能性があります。 そして、抗原と抗体に対する耐性の軸は何によっても分離されていません。
化学的攻撃は、タンパク質分子のさまざまな毒素への結合に影響を与えるため、体は安全に解毒することができます。 私たちの体の解毒の最も重要な役割は、ほこりを分解してそれらをrozchinnuの形に変換する肝臓酵素によって果たされます。 rozchinenіtoxiciswidkoは生物を枯渇させます。
規制機能
内部の細胞プロセスのほとんどは、タンパク質分子によって調節されています。 気分子vykonuyut高度に専門化された機能、およびєbudіvelnymkіtinnym材料、nіdzherelエネルギー。 調節は、酵素の活性と他の分子に結合する酵素の影響を受けます。中細胞におけるプロセスの調節の重要な役割は、プロテインキナーゼによって果たされています。 これらは、リン酸粒子を追加することで他のタンパク質の活性を追加する酵素です。 悪臭の気は活動を強化し、気はїїを窒息させます。
信号機能
白人の信号機能は、信号スピーチとして機能するように建物に現れています。 悪臭は、組織、細胞、臓器の間で信号を送信します。 いくつかのシグナル伝達機能は、調節機能と同様に重要ですが、豊富に調節される内部細胞タンパク質の断片もシグナル伝達に影響を与えます。 細胞は、細胞間発話を通じて拡大する追加のシグナルタンパク質のために互いに相互作用します。サイトカイン、タンパク質-シグナル伝達機能のためのホルモン。
ホルモンは血液によって運ばれます。 受容体は、ホルモンに結合すると、クリチナムで反応を引き起こします。 ホルモンは、血球の発話濃度の調節、および細胞の成長と生殖の調節に影響を及ぼします。 このようなタンパク質のお尻は、血中のブドウ糖の濃度を調節する広く使用されているインスリンです。
サイトカインは小さな情報ペプチド分子です。 悪臭は、異なるクリチン間の相互作用の調節因子として機能し、これらのクリチンの生存を意味し、それらの成長と機能的活動を抑制または刺激します。 サイトカインがなければ、神経系、内分泌系、免疫系の機能は不可能です。 たとえば、サイトカインは腫れの壊死を引き起こす可能性があります。つまり、炎症細胞の成長と活力が影響を受けます。
トランスポート機能
残留タンパク質は、小分子の輸送に関与するため、高濃度であるため基質に容易に結合し、低濃度で見つかるとそこで振動する原因にもなります。 輸送タンパク質のお尻はヘモグロビンです。 それは肺から酸素を輸送して他の組織に運び、またそれを組織から軽い二酸化炭素に逆方向に輸送します。 生物のすべての王国で、ヘモグロビンに類似したタンパク質が知られていました。予約(または予約)機能
このようなタンパク質には、カゼイン、卵白アルブミンなどが含まれます。 動物の卵細胞や在来植物の気予備タンパク質は、エネルギー源として貯蔵されています。 悪臭は生命機能を殺している。 多くのタンパク質は、多くのアミノ酸のように私たちの体に見られます。タンパク質の受容体機能
タンパク質受容体は、細胞膜と細胞質の両方に存在する可能性があります。 タンパク質分子の一部が信号を受信します(性質が化学、光、熱、機械のいずれであっても)。 タンパク質受容体は、入ってくる信号に応答してコンフォメーション変化を認識します。 他の細胞成分の信号伝達を担当するため、分子の他の部分を追加するための変更。 信号の伝達メカニズムは1つずつ異なります。モーター(または手)機能
モータータンパク質は、(体のレベルでの)m'yazyvの循環とスキャリーの安全性、およびべん毛とviyの動き、発話の内部細胞輸送、白血球のアメーバ増殖(生物のレベルで)に関与しています。 )。スピーチの交換におけるタンパク質
現場のロスリンと微生物のほとんどは、20の基本的なものと、追加のアミノ酸のスプラットを合成します。 しかし、なんて臭い dovkilla、それから生物はエネルギーを節約し、それを真ん中で輸送し、それを合成しません。体内で合成されないアミノ酸は必須と呼ばれているため、私たちのところに来ることはあまりありません。
人々は静かなタンパク質からアミノ酸を取り除いて、ハリネズミから取り除くことができるようにします。 タンパク質は、酸っぱい殻のジュースや酵素の影響下で、エッチングの過程で変性します。 アミノ酸をエッチングするプロセスの結果として、それらの一部は必要なタンパク質の合成のために停滞し、糖新生のプロセスの一部はグルコースに変換されるか、クレブス回路(代謝崩壊のプロセス)で停滞します。
エネルギー源としてのVikoristannyaタンパク質は、体に内部の「不十分な供給」、つまり水タンパク質がある場合、不親切な心の中で特に重要です。 体のアミノ酸も重要な窒素源です。
タンパク質の追加消費に関する統一された基準はありません。 腸に生息する微生物叢もアミノ酸を合成し、悪臭はタンパク質基準の時間の下で保護することができます。
人体のたんぱく質の貯蔵は最小限であり、新しいたんぱく質は、分解されたたんぱく質、体の組織、そして一緒に見つかるアミノ酸からのみ合成することができます。 脂肪と炭水化物の倉庫に入るサイレントスピーチから、タンパク質は合成されません。
行方不明のリス
食事中のタンパク質の発話の欠如は、子供の発達の発達のより強い増加につながります。 高齢者の場合、肝臓の深い変化の出現、ホルモンの背景の変化、内分泌の機能障害、生きている言語の習得の低下、記憶と練習の障害、心臓の問題のために、タンパク質の欠乏は安全ではありません。 これらすべての否定的な症状は、タンパク質が人体のすべてのプロセスに関与している可能性があるという事実によるものです。
前世紀の70年間に、致命的な噴火が、ほとんどの場合、顕著なタンパク質欠乏を伴う低カロリーの食事をしている人々で記録されました。 原則として、この場合、心臓の肉の取り返しのつかない変化が、媒介されない死因でした。
タンパク質の欠乏は感染症に対する免疫力を低下させ、抗体産生のレベルが変化します。 インターフェロンとリゾチーム(抗酸化因子)の合成障害は、発火プロセスの強化を引き起こします。 さらに、タンパク質の欠乏はしばしばビタミンの不足を伴い、それはまた不利な結果につながります。
欠乏は、酵素の生成や重要な生きたスピーチの同化にはもはや寄与しません。 タンパク質溶液を含むホルモンも、少量のタンパク質が深刻なホルモン破壊を引き起こす可能性があることを忘れないでください。
それが肉体的な性格の活動であろうと、shkodi m'azovim clitinのリーダーであり、虚栄心が高ければ高いほど、m'yaziはより苦しみます。 貧しいクリチン・マヤジブの復活には、大量のヤコウタンパク質が必要です。 心を広げてみると、十分な量のたんぱく質が体に供給されていれば、肉体的な野心は同じ色よりも少なくなります。 激しい物理的圧力により、タンパク質の減少は、皮膚のキログラムあたり皮膚の1.5〜2グラムに達する可能性があります。
タンパク質が多すぎる
体内の窒素バランスを維持するためには、一定量のタンパク質が必要です。 食事に含まれるタンパク質が多い場合は、タンパク質が3つ以上あるため、健康に害を及ぼすことはありません。 私の意見では、補助エネルギーのような世界的なアミノ酸の数vikoristovuєtsya。しかし、スポーツに参加せず、水1キログラムあたり1.75 gを超えるタンパク質を摂取している場合、肝臓に蓄積されるタンパク質が多すぎて、窒素含有量とブドウ糖に変換されます。 Azotiste z'ednannya(sechovin)は、obov'yazkovo vyvoditsyanirkamiz生物である可能性があります。
さらに、ワインのタンパク質が過剰になると、体の反応が酸っぱくなり、食事療法の変更を通じてカルシウムが消費されます。 それまでは、たんぱく質が豊富な肉用ハリネズミは、代謝過程にあるものなど、しばしばプリンを復讐し、沼地に堆積し、痛風の発症を呼びかけます。 重要なことに、過剰なタンパク質に関連する損傷は、タンパク質の欠乏に関連する、より豊富でより低い損傷をトラップします。
食事中の十分な量のタンパク質の評価は、窒素バランスキャンプによって決定されます。 生物では、新しいタンパク質の合成とタンパク質代謝の最終産物の名前の出現が継続的に行われています。 窒素はタンパク質貯蔵庫に入りますが、これは脂肪にも炭水化物にも見られません。 そして、窒素は予備として体内に貯蔵されているので、ビルキブの倉庫にのみ貯蔵されています。 静脈のタンパク質崩壊の場合、セクションから名前をすぐに見たのは責任です。 生物の機能が必要なレベルで実行されるためには、見られる窒素を補充する必要があります。 窒素バランスとは、還元される窒素の量が体内に導入される窒素の量に等しいことを意味します。
Bilkoveを食べる

幼虫タンパク質のクラストは、タンパク質吸収係数によって推定されます。 この係数は、化学的値(アミノ酸の倉庫)と生物学的値(タンパク質の過剰摂取量)です。 これらの製品に含まれるタンパク質の全量を使用すると、同化係数が1.00に等しくなる可能性があります。
卵、大豆たんぱく質、牛乳などの製品のゲイン係数は1.00です。 Yalovychynaは0.92の係数を示しています。
Qi製品єvysokoyakіsnyjdzherelomタンパク質、タンパク質の痕跡記憶、学校の復讐は脂肪が豊富で、食事療法の頻度を害する必要はありません。 クリームは大量のタンパク質であり、体内では世界中の脂肪を消費する必要もあります。
たんぱく質含有量が豊富な優れた製品:大豆シリ、低脂肪シリ、低脂肪子牛肉、卵白、低脂肪チーズ、新鮮な魚介類、若い子羊、鶏肉、白身の肉。
それほど重要ではないのは、ズクルを加えたミルクとヨーグルト、赤身の肉(ビリズカ)、ダークチキンとインドの肉、低脂肪のナリズカ、自家製チーズ、ベーコン入りの加工肉、サラミ、シンキなどです。
卵白-脂肪がない純粋なタンパク質。 犬の肉はカロリーの約50%を含んでいるため、タンパク質が追加されています。 でんぷんを含む製品の場合-15%; 低脂肪乳で-40%; 野菜で-30%。
プロテインミールを選ぶときのスマットルールは攻撃的です: 多数単位カロリーあたりのタンパク質と高いタンパク質吸収係数。 低脂肪と高タンパク質の製品を使用するのが最善です。 カロリーに関するデータは、どの製品のパッケージにも記載されています。 これらの製品に含まれるタンパク質と脂肪の量に関するより詳細なデータは、カロリー量を簡単に差し引くことができ、特別な表に記載されています。
熱処理を認識したタンパク質を征服するのは簡単で、悪臭の破片がハーブ管に酵素を注入できるようになります。 ただし、温度処理は、アミノ酸を脱酸するものを介してタンパク質の生物学的価値を低下させる可能性があります。
Vmіstblіkіvizhirivvdeyakyhgrub製品
| 製品 | タンパク質、グラム | 脂肪、グラム |
| 鶏 | 20,8 | 8,9 |
| 心臓 | 15 | 3 |
| 豚肉は痩せています | 16,3 | 27,8 |
| Yalovychyna | 18,9 | 12,3 |
| 子牛の肉 | 19,7 | 1,2 |
| ドクトロスカヤヴァレナカウバス | 13,7 | 22,9 |
| ディーティクナ茹でカウバス | 12,2 | 13,5 |
| ミンタイ | 15,8 | 0,7 |
| オセレーデツィ | 17,7 | 19,6 |
| チョウザメのキャビア粒状 | 28,6 | 9,8 |
| ボロスナIグレードの小麦パン | 7,6 | 2,3 |
| ライフパン | 4,5 | 0,8 |
| 良いvipіchka | 7,2 | 4,3 |
ミルクに含まれるカゼインもトッププロテインです。 新しい道路での取得係数1.00。 牛乳から見ると、カゼインと大豆は、乳糖不耐症に苦しむ人々が生きることができるように、悪臭が乳糖を復讐しない、タンパク質含有量の高い角質食品を作る機会を与えます。 そのような製品のもう一つの利点は、アレルゲンの潜在的な供給源として、それらがsyrowortsを含まないことです。
タンパク質代謝

タンパク質を獲得するために、体は多くのエネルギーを必要とします。 私たちの生物は、タンパク質のアミノ酸ランスを短いランスのスプラット、またはアミノ酸自体に分割することで罪を犯しています。 このプロセスは、体がハーブ管の作成と輸送に責任があるため、トリバリウムを追加し、さまざまな酵素を助けることです。 タンパク質代謝の過剰な生成物である窒素のスラッグは、体から取り除くことができます。
合計spozhivayutchimaluのすべてのtsіpodіїは、タンパク質の獲得のために多くのエネルギーを持っています。 したがって、タンパク質は代謝の加速と内部プロセスのエネルギーレベルの増加を刺激します。
生物の同化については、食事のカロリー量にもよりますが、15%近くを費やすことができます。
代謝の過程でタンパク質の含有量が高いため、それは熱生成を増加させます。 トロッチの体の温度が上昇し、それが熱発生のためのエネルギーの追加の増加につながります。
タンパク質は必ずしもエネルギー物質のようにビコレートするとは限りません。 これは、体のエネルギー源としてのzastosuvannaїхが見えなくなるという事実によるものです。大量の脂肪と炭水化物があっても、より多くのカロリーをより効率的に摂取し、同じ量のタンパク質を減らすことができます。 その前は、体内に過剰なタンパク質が存在することはめったになく、さらに多くのワインが存在する場合、可塑性機能の発達のために体内に過剰なタンパク質が存在します。
その場合、食事にエネルギー源がなければ、脂肪と炭水化物を見ることができ、体はより多くの蓄積された脂肪のために取られます。
食事中の十分な量のタンパク質は、肥満に苦しんでいる静かな人々の発話の代謝の増加を活性化および正常化するのに役立ち、また、m'yazovuの質量を改善することができます。
たんぱく質がくっつかない場合、体は勝利したm'yazovykhたんぱく質と混ざり合っています。 m'yaziは生物の生命を支えるのに大したことではないという事実に注意してください。 m'yazovyh繊維では、より多くのカロリーが燃焼され、m'yazovyの質量が減少し、体のエネルギー消費が減少します。
さらに多くの場合、減量のために別の食事をとろうとしている人々は、そのような食事を選択します。その場合、体内のタンパク質はほとんどありません。 原則として、野菜や果物の食事。 クリムシュコディ、そのような食事は何ももたらさないでしょう。 タンパク質が不足している場合の臓器系の機能を考慮に入れる必要があり、それはさまざまな損傷や病気につながります。 皮膚の食事は一目で見て、タンパク質から体が消費する必要があります。
タンパク質の同化とエネルギー需要におけるそれらの維持、およびタンパク質代謝産物の導入などのプロセスには、より多くのリンジンが必要になります。 水を奪わないために、当日は2リットル近くの水を飲む必要があります。
膜貫通型イオンストラム。
受容体に特異的に結合する発話は、その受容体のリガンドと呼ばれます。 体の真ん中で、ホルモン、神経伝達物質、または代替品のいずれかを呼び出します。これは、医薬品およびアルコール(アゴニスト)としてzastosovuyutsyaです。 Deyakіligandi、navpaki、ブロック受容体(拮抗薬)。 器官の感覚、発話のリガンドがある場合は、受容体に香りを注入するか、味わってください。 さらに、口腔受容体の分子は光に反応し、聴覚器官や点状器官では、受容体は刺し傷や他のサブかみそりによって呼ばれる機械的注入(圧力または張力)に敏感です。 膜電位の変化に反応する感熱性受容体タンパク質と受容体タンパク質もあります。
百科事典YouTube
-
1 / 5
クリチン受容体膜受容体と細胞内受容体の2つの主要なクラスに分けることができます。
膜受容体
「アンテナ」の機能は、音声信号の認識です。 Dіlyankidvohsusіdnіhkіtin、shchorazznіyutは、クリチンの接続を保護し、1対1でリンクすることができます。 Zavdyaki tsymu clitinyは方向付けられ、分化の過程で組織を作成します。 認識可能な細胞が存在し、さまざまな方法で知られているさまざまな分子で、悪臭がクリチンによって振動的に粘土化されている理由は、認識されている相補的な細胞である可能性があります(たとえば、LPGはLDL受容体の助けを借りて崇拝されています)。
膜受容体の2つの主要なクラスは、代謝型受容体とイオノトロピック受容体です。
イオノトロピック受容体は、リガンドに結合すると開閉する膜チャネルです。 Ionni Strumi、TsomoのVinikayat、ViclicayuZmіni膜貫通型RIZNITSIPotzіalivI、vnaslіdokTsooi、Zbudno -Klitini、そして彼は内部濃縮物のヘビであり、彼らは問い合わせの評価に生き残ることができます。 最近開発されたイオノトロピック受容体の1つは、n-コリン作動性受容体です。
代謝型受容体は、内部細胞メディエーターのシステムによって結合されます。 リガンドと結合したときにそれらのコンフォメーションを変化させて、生化学反応のカスケードを開始させ、おそらく細胞の機能状態を変化させます。 膜受容体の主な種類:
- ヘテロ三量体Gタンパク質に結合する受容体(たとえば、バソプレシン受容体)。
- 内因性チロシンキナーゼ活性を促進する受容体(例えば、インスリン受容体または上皮成長因子受容体)。
Gタンパク質、7つの膜貫通ドメイン、N末端および細胞内C末端を含む膜貫通タンパク質にリンクされた受容体。 リガンドとの結合部位は亜クリチナスループ上にあり、Gタンパク質との結合ドメインは細胞質のC末端近くにあります。
受容体の活性化は、αサブユニットがβγサブユニット複合体に解離して活性化される前に実行されます。 もしそうなら、それは酵素を活性化するか、さもなければ不活性化し、二次中間体を生成します。
チロシンキナーゼ活性を有する受容体は、しばしばプロテインキナーゼによっても開始細胞内タンパク質をリン酸化し、このようにしてシグナルをクリチンの内部に伝達する。 構造によると、それらは1つの膜ドメインを持つ膜貫通タンパク質です。 原則として、ホモ二量体、そのようなコーティングのサブユニットは、ジスルフィドパッチで覆われています。
細胞内受容体
細胞内受容体-原則として、転写因子(例えば、糖質コルチコイド受容体)または転写因子と相互作用するタンパク質。 内部細胞受容体のほとんどは、細胞質内のリガンドと結合し、活性状態に移行し、リガンドと一緒に細胞の核に輸送され、そこでDNAに結合し、活性遺伝子またはグループの発現を誘導または抑制します。遺伝子の。
一酸化窒素(NO)の形成のための特別なメカニズム。 膜を透過するこのホルモンは、細胞質ゾルのグアニル酸シクラーゼ、一酸化窒素受容体、および二次中間体であるcGMPを合成する酵素に結合します。ホルモンシグナルの細胞内伝達の主なシステム
アデニル酸シクラーゼシステム
アデニル酸シクラーゼシステムの中心部分は、ATPからcAMPへの変換を触媒する酵素アデニル酸シクラーゼです。 この酵素は、G sタンパク質(英語刺激下)によって刺激されるか、G iタンパク質(英語抑制下)によって無視されます。 次に、cAMPはcAMPが枯渇したプロテインキナーゼ、いわゆるプロテインキナーゼA、PKAに結合します。 タンパク質エフェクターのその遠いリン酸化の活性化を引き起こす必要があり、それはクリチンにおいて生理学的役割を果たすでしょう。
ホスホリパーゼ-カルシウム系
G qタンパク質はホスホリパーゼ酵素を活性化し、PIP2(膜ホスホイノシトール)をイノシトール-3-リン酸(IP3)とジアシルグリセリドの2つの分子に分割します。 これらの分子の皮膚は2番目の中間体です。 IP3は、小胞体の膜上の受容体に結合することができます。これにより、カルシウムが細胞質に放出され、さまざまな細胞反応が開始されます。
グアニル酸シクラーゼシステム
このシステムの中心分子はグアニル酸シクラーゼであり、GTPからcGMPへの変換を触媒します。 cGMPは、低酵素とイオンチャネルの活性を調節します。 グアニル酸シクラーゼのІsnuєスプラットアイソフォーム。 それらの1つは、心房性ナトリウム利尿因子受容体に間接的に結合している一酸化窒素によって活性化されます。
cGMP制御交換は、腸と腸でのイオン輸送につながり、心臓ではリラックスのシグナルとして機能します。
受容体の薬理学
原則として、健康な受容体は、主要な内因性リガンドだけでなく、他の構造的に類似した分子にも結合します。 この事実により、受容体と結合し、顔や拭き取りなどの状態を変化させる、vikoristovuvatyの外因性発話が可能になります。
したがって、たとえば、エンドルフィンの受容体-痛みや感情状態の調節に重要な役割を果たす神経ペプチドは、モルヒネグループの薬と関連しています。 受容体は、母体、主要部位のクリム、またはその受容体に特異的なホルモンとのコミュニケーションの「部位」、またはメディエーター、ならびに他の発話化学物質が接続されている追加のアロステリック調節部位である可能性があります。ホルモン受容体シグナルは(変化)に基づいています-ブーストまたは弱体化、またはメインシグナルを置き換えます。 さまざまな発話に結合するデシルコマのそのような受容体の典型的な例は、受容体-γ-アミノ酪酸A型(GABA)です。 GABA自体のリンクサイトに加えて、ベンゾジアゼピンのリンクサイト(「ベンゾジアゼピンサイト」)、バルビツール酸塩のリンクサイト(「バルビツール酸塩サイト」)、アロプレグネノロンタイプの神経ステロイドのリンクサイト(「ステロイドウェブサイト」)もあります。 ")。
バガト型受容体は、ibyііivvyazuvannya kilka ryznikh nyznikh nymichychnyから出ることができます、私は特定の川の出発で、彼女はノックアウトしました、構成の繊維のいとこを知っています-「割り当てられていません)(そこに)受容体にはホルモンがありません)が、いくつかの中間体にもあります。
受容体に結合すると、受容体を「100%オン」構成に切り替える、100%の免疫を備えた音声は、受容体の最終アゴニストと呼ばれます。 スピーチ、zі100%ymovіrnіstyu、vyklykaє受容体と結合するとき、yogoの構成「100%ワクチン接種」への移行は、受容体の逆アゴニストと呼ばれます。 受容体の中間構成の1つへの移行、または受容体の変化を要求するスピーチは、100%変化しません(そのため、受容体の一部は、スピーチに接続されたときに、オンまたはオフになり、一部受容体のはしばしば駆動されます)。 そのようなスピーチを参照することにより、アゴニスト-アンタゴニストという用語が使用されます。 結合時に受容体を変化させず、ホルモン受容体またはメディエーターへの結合をあまり受動的に伝達しないスピーチは、競合的拮抗薬または受容体遮断薬と呼ばれます)。
原則として、外因性の発話は体の中央に受容体を持っていますが、体の中にはこの受容体の内因性のリガンドがあります。 したがって、たとえば、ベンゾジアゼピンの内因性リガンド