Інніチャネルとїхbudova。 生体膜およびイオンチャネル。 イオンチャネルの構造と機能に関する重要な啓示
イオンチャネルの力
選択性-cevibirkovaは、イオンを歌うためのICの浸透を増加させました。 他のイオンの場合、浸透が減少します。 このような活気は、チャネルの多孔性の最も重要な部分である選択フィルターによるものです。 フィルター、ナローカットのクリーム、おそらく地元の 電荷。 たとえば、陽イオン選択チャネルは、タンパク質分子の倉庫内の負に帯電した過剰なアミノ酸を選択フィルターの領域で鳴らすため、正の陽イオンを引き付け、負の陰イオンを放出し、細孔を通過しません。
Kerovanaの浸透-ІХvіdkrivatisyaの構築、またはチャンネルで歌うkeruchihvplivіvのために丸くなります。 閉じたチャネルは浸透を減らし、開いたチャネルは増加する可能性があります。 電力のために、ICはさまざまな方法で休耕地に分類できます。たとえば、電位活性化、リガンド活性化などです。
不活性化-開封後1日以内にІHを構築する目的は、それが振動し、機能し続ける活性化要因である場合、その浸透をポイントまで自動的に減らします。 Shvidkaの非アクティブ化は、チャネルが適切に閉じられた場合に実行される独自の特別なメカニズムを備えた特別なプロセスです(一般的な非アクティブ化)。 プロセスと並行してプロセスを処理するには、チャネルを閉じる(完全に非アクティブ化する)必要があります。これにより、プロセスは必要な情報を提供します。 rahunokはチャネルタンパク質のコンフォメーションを変更します。 そして、軸は、例えば、電位活性化チャネルで、スウィドカ不活性化は、まるで風呂でバイコリストのように、ランセットのコルクを推測する特別な分子「コルクシール」の助けを借りて使用されます。 このプラグはアミノ酸(ポリペプチド)ループで、最後に汗があります 3つ見て zakaetsyavnutrishnegirloが細胞質の側面からチャネルするアミノ酸。 Саме тому потенціал-залежні ІЧ для натрію, що забезпечують розвиток потенціалу дії та рух нервового імпульсу, можуть пропускати в клітину іони натрію тільки протягом декількох мілісекунд, а потім вони автоматично закриваються своїми молекулярними пробками, незважаючи на те, що деполяризація, що відкриває, продовжує子供。 ICの不活性化の別のメカニズムは、追加のサブユニットによる内側のクリチナスチャネルの変更である可能性があります。
ブロッキングは、キャンプの1つとして修正し、zvichaynіkeruuchііїに反応しないように、スピーチブロッカーのІChpіddієyuを構築することの目標です。 このような状態では、チャネルは単に重要なメッセージに関するフィードバックを提供しなくなります。 ブロッキングは、敵対者、ブロッカー、またはブロッカーと呼ばれる可能性のあるスピーチブロッカーの呼び出しをブロックします。 拮抗薬-他のスピーチの活性化をICにシフトするスピーチライン。 したがって、建物のスピーチは受容体セルІChとよく関連していますが、建物ではなく、ステーションをチャネルに変更し、音声の応答を変更します。 受容体の封鎖をすぐに終了します-ІChの封鎖。 拮抗作用は受容体とyogoІChの同じ封鎖を引き起こさないことを覚えておいてください。悪臭はロボットチャネルに対して弱くなり、ingibuvati(prignichuvati)が少なくなる可能性がありますが、それを再び非難することはできません。 作動薬-拮抗薬-ceスピーチ、yakіは受容体に弱い刺激注射をするかもしれませんが、同時に、制御されるべき自然の内因性レコビナの発達をブロックします。 ブロッカー-ロボットのイオンチャネルを通過するサービス、たとえば、メディエーターと相互作用して新しい分子受容体への分子受容体と相互作用することも、それらがブロックするチャネルの制御を妨害します。 たとえば、抗コリン作用薬は地獄のアセチルコリンを遮断します。 アドレナリンを含むノルエピネフリン-アドレナリン遮断薬; ヒスタミン-ヒスタミン遮断薬など。多くの遮断薬は、治療目的で医薬品として使用されます。 溶解物はそれ自体がブロッカーであり、この用語はブロッカーの同義語としてより古く、勝利を収めています:コリン分解、アドレナリン分解など。
可塑性は、その力、その特性を構築し、変更することの価値です。 可塑性を保証するメカニズムの最大の拡張は、プロテインキナーゼ酵素による膜の内側からのチャネルタンパク質のアミノ酸のリン酸化です。 ATPとGTPのリン酸の余剰分がチャネルタンパク質に追加され、チャネルはその力を変化させます。 たとえば、閉じた状態で固定されたり、逆に開いた状態で固定されたりします。
ブドフとイオンチャネルの機能。 それらはNa+、K +、Ca 2+、Cl-セルの中央に浸透し、特別な方法で名前を入力し、マザーチャネルを埋めます。 Rozmir kanaliv dosit maly(直径05-07 nm)。 Rosrahunkiは、運河の総面積が表面のわずかな部分を占めていることを示しています 細胞膜.
イオンチャネルの機能 別の方法で。 最も拡張してみましょう-電圧を固定する方法、または「電圧クランプ」(図2.2)。 この方法の本質は、特別な電子システムの助けを借りて、プロセスが膜電位を同じレベルに変更および固定できることです。 これにより、膜を流れるイオン性ストラムの大きさが制御されます。 電位差がになっている場合、オームの法則によれば、ストリームの値はイオンチャネルの導電率に比例します。 脱分極の同じ段階で、電気化学的勾配に沿ってクリチンに入る他のチャネル、すなわち、クリチンを脱分極するイオン性ストラムがあります。 Qiaの変更は、keruyuchy podsiluvachの助けを借りて登録され、膜を通過します 電気ストラム、大きさに等しいが、膜イオン性ストラムの直線で増殖している。この場合、膜貫通電位差は変化しない。 Spіlnevikoristannyaイオンチャネルの潜在的かつ特定のブロッカーを固定する方法が承認につながった 他の種類細胞膜のイオンチャネル。
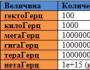
このとき、さまざまなイオン用のさまざまなタイプのチャネルが設置されています(表2.1)。 それらのいくつかはより具体的であり、他のものは、メインイオンを除いて、他のイオンをスキップすることができます。
パスクランプ電位への局所固定の方法による、可能な限り多くのチャネルの機能の確立。 米。 2.3 A)。 微小電極(マイクロピペット)を塩水噴霧で絞り、膜の表面に押し付けて小さな噴霧を作成します。 同時に、膜の一部が微小電極まで移動します。 ピックアップゾーンでイオンチャネルが検出されるとすぐに、単一チャネルのアクティビティが登録されます。 チャネルへの活動のいじめと登録のシステムは、電圧を固定するシステムの影響をほとんど受けません。
表2.1。警戒心の強い人々の最も重要なイオンチャネルとイオンストリーム
ノート。 TEA、テトラエチルアンモニウム; TTX-テトロドトキシン。
運河の外側は部分的にアクセス可能ですが、運河の内側は重要になります。 P. G. Kostyukは、微小電極をブロックすることなく、イオンチャネルの入口および出口構造の機能を可能にする細胞内透析の方法を開発しました。 喉のスペースの後ろで開いているイオンチャネルの一部は、その機能的な力のために、チャネルの一部で乾燥し、最も内側の中央で動物化されているように見えました。
イオンチャネル自体は、選択性と導電性という2つの重要な膜力を提供します。
選択性、また 活気、チャネルは特別なタンパク質構造によって保護されています。 膜電位の大きさに応じて、これらの建物を堆積させることができるように、多数のチャネルが帯電しています。 チャネルは、その機能的特徴、特にチャネルへの入口とその出口(いわゆるcomirn_mechanisms)として知られているタンパク質構造の重要性に関して不均一です。
5.起きていることについて理解する。 神経-舌系の警戒のパラメータ:からかいの閾値(レオベース)、朝の時間(クロナキシー)。 1日の時間におけるrozdratuvannyaの強さの休耕地(Gorweg-Weiss曲線)。 耐火物。
覚醒-razdratuvannya成形PDと特定の反応に関するZdatnіstkіtinyvіdpovіdati。
1)局所脱分極の段階-膜の頻繁な脱分極(クリチンへのNa +の流入)。 小さなドレッサーを塗ると丈夫になります。
局所脱分極-高揚期。
2)絶対的な不応性の段階-どんな種類の力でもPDを形成しない覚醒組織の力
3)視覚的不応性の段階。
4)完全な再分極の段階-razdratuvannya-再び強い再分極
5)過分極の段階-注意力が低い(正常以下)場合、刺激は大きくなる可能性があります。
機能不安定性-1時間で可能な最大数のPDによる組織の注意力の評価。
ダメージを止めます:
1)強さの法則-podraznikの強さは、しきい値またはしきい値を超えることができます(覚醒を呼びかける力の最小値)。 最強のいじめ、最強の目覚めは何ですか-組織の凝集が少ない(神経質なstovbur、m'yaz、vinyatok-MMC)。
2)時間の法則-些細な衰弱のpodraznikは、有罪の目覚めの呼びかけには十分である可能性があります。
その1時間前の力によるミズ、最小時間によるミズの境界での比例休閑と最小力によるそれ。 最小の強さ-レオベース-は、目覚めと風に横たわることを要求する強さです。 最小時間はブルーアワーです。 クロナキシー-目覚めの原因であるこれらの他の組織の警戒心、1時間は、2回の再観察を行います。
強さが大きければ大きいほど、歌の価値に対する強さが大きくなります。
WFPを作成する職員:
1)ナトリウムとカリウムの濃度の違い
2)ナトリウムとカリウムの浸透率
3)Na-Kポンプの動作(3つのNa +がポンプで排出され、2つのK +が回転します)。
生きている構造物の反応の最小の反応に必要な、podraznikの強さとその三位一体の間の休閑は、いわゆる力曲線-時間(Goorweg-Weiss-Lapik曲線)を歩く方が良いです)。
曲線の分析から、podraznikの強度が非常に大きく、三位一体が不十分であっても、流入に反応がないことは明らかです(誇張の粘性のある首のレボルフのポイント)。 同様の現象は、trivalo disubthresholdpodraznikiでも保護されています。 ストラム(または張力)の最小強度であるzdatna viklikati zbudzhennyaは、Lapik rheobase(vіdrіzok縦方向OA)によって命名されました。 ストラムのように伸び、目覚めの構造で呼びかけられる、レオバシーの底流の強さに等しい1時間の最小間隔は、クロナキシー(OFの横座標)と呼ばれ、しきい値の三位一体の指標として使用されます。からかいの。 クロナキシーはδ(1000分の1秒)減少します。 クロナキシーの大きさは、ファブリックの覚醒の重症度を判断するために使用できます。クロナキシーが少ないほど、覚醒が高くなります。 人間の神経と粘膜の繊維のクロナキシーは1000分の1秒と古く、ヒキガエルの嚢の粘膜繊維などの主要な組織のクロナキシーは100分の1秒です。
覚醒している組織の慢性化は、実験において、そしてスポーツの生理学において、診療所において広範囲に及ぶ。 クロナキシー・マヤザの緩和の道の助けを借りて、神経病理学者は、ルーチ神経の破裂の存在を確立することができます。 弱者を強くすることができ、母親は境界線の三位一体であることを示す必要がありますが、低速は1時間で境界線の値まで増加します。その場合、それを非難することはありません。 異常増殖の全範囲への覚醒組織の付着は、宿泊施設の名前を取り除いた。 宿泊施設は、1時間で生地の下着の強度の成長が上がるという事実によって揺らいでいます。 アクティブな変更、からかいのしきい値を上げ、目覚めの発達を再シャッフルするこのような儀式では、目覚めの目覚めには、時間でのからかいの成長の速度、またはからかいの勾配が重要です。
勾配開発の法則。 ティーザーへのライブ照明の反応は、からかいの勾配の形で落ちることです。これは、時間内のティーザーの成長の用語または涼しさによるものです。からかいの最大の勾配は何ですか、これは目覚めている照明の空気の中で最も強い(歌うインターへの)反応。
同じ法則から、いじめは、їїvzaєmodіїのティーザーとアウェイク構造の間で相互に折りたたまれる必要があります。 viniknennya zbudzhennya podraznikは、母親の国境の強さ、母親の国境の三位一体、母親のpevnuの急成長の罪を犯しています。
6.イオンポンプ(ATP-azy):K + -Na + -eva、Ca2 + -eva(筋小胞体のプラズモレマイト)、H + –K+交換体。
最近の発見によると、生体膜には、ATP加水分解の自由エネルギーのために働くイオンポンプがあります-内在性タンパク質(輸送ATPアーゼ)の特別なシステム。
現在、膜を介してイオンを活発に移動させる起電性イオンポンプには3つのタイプがあります(図13)。
輸送ATPアーゼによるイオンの移動は、クリチンのエネルギー代謝のための化学反応を伴うその後の移動プロセスによるものです。
皮膚ATP分子の加水分解中に発生するエネルギー変動に対するK+-Na + -ATPaseの作業中に、2つのカリウムイオンがクリチンに移動し、3つのナトリウムイオンが同時にクリチンから排出されます。 このようにして、クリチニウム中のカリウムイオンの濃度が減少します。これは生理学的に非常に重要です。
「バイオポンプ」の兆候:
1.電気化学ポテンシャルの勾配に逆らって急いでください。
2. ATP(または他のエネルギー源)の加水分解からの包帯のスピーチの流れ。
3.輸送機の非対称性。
4. ATPのポンプinvitro構築加水分解は、invivoで移動できる静かなイオンの存在下では少なくなります。
5.ポンプが一体に取り付けられている場合、静脈の中央で選択性を節約できます。
ロボットATPアーゼの分子メカニズムは未解決のままです。 折り畳み酵素プロセスの主な段階は、プロテグされています。 さまざまなK+-Na + -ATフェーズでは、ATP加水分解によるイオン移動のシミュレーション段階があります。
この図は、酵素の働きにおける重要なステップが次のとおりであることを示しています。
1)膜の内面でのATPとの酵素複合体の固定(反応はマグネシウムイオンによって活性化されます);
2)トリオキソイオン性ナトリウム錯体との結合;
3)アデノシン二リン酸の変換のための酵素のリン酸化;
4)膜の真ん中で酵素にフリップ(フリップフロップ)します。
5)膜の外面で反応するカリウムのナトリウムイオン交換の反応。
6)カリウムイオンが細胞の中央に移動することによる酵素複合体の逆転。
7)酵素をに変える 休日のキャンプカリウムイオンと無機リン酸塩(P)のレベルに。
このようにして、最後のサイクルで、ナトリウムの3つのイオンが細胞から放出され、2つのカリウムイオンが細胞質で濃縮され、1つのATP分子が加水分解されます。
これらのチャネルは、オリゴマー(デカールサブユニットで構成される)タンパク質である細胞膜で特に確立されます。 タンパク質分子は、親水性の中心にチャネル細孔が形成されるように膜を貫通し、スリットを通って建物のクリチナに浸透するため、中央チャネルへのタンパク質分子であり、その直径は細孔の直径を超えないようにしてください(音tseioni)。
チャネルのヘッドサブユニットの近くで、デカールサブユニットのシステムが開発され、膜調節タンパク質、さまざまなメディエーター、および薬理学的に活発なスピーチとの相互作用のためのプレートを形成します。
それらの機能のためのイオンチャネルの分類:
1)チャネルが透過性であるイオンの数について、チャネルは選択的(1種類のイオンのみを透過)と非選択的(デカール種のイオンを透過)に分けられます。
2)イオンの性質に応じて、Na +、Ca ++、Cl-、K+-チャネルを通過させるためにヤキ悪臭を放ちます。
3)規制の方法に従って、潜在的な預金と潜在的な独立に分けられます。 電位依存チャネルは、クリチン膜の電位の変化に反応し、歌の値の到達可能な電位を超えて、チャネルはアクティブステーションに変わり、濃度勾配から徐々にイオンを通過させます。 したがって、ナトリウムチャネルとカルシウムチャネルは電位が沈着し、膜電位が-50〜60 mVに低下したときに活性化が起こり、血液中のNa+およびCa++イオンの流れが穏やかな遺伝子の電位を低下させます。 カリウム沈着チャネルは、PD Iの発生中に活性化され、細胞からのK +イオンのストラムを防ぎ、膜の再分極を引き起こします。
電位に依存しないチャネルは、膜電位の変化に対して異なる反応を示しますが、相互作用の匂いとリガンドの匂いのために、受容体の相互作用に対しては反応します。 したがって、Clチャネルはg-アミノ酪酸受容体と結合し、これらの悪臭受容体が相互作用すると、それらは活性化されて塩素イオンの流れをクリチナに提供し、過分極と覚醒の低下を引き起こします。
3.安心のための膜電位。
期間 「膜電位心の平和 » 細胞質と最も一般的な細胞の違いの間にある、電位の膜貫通差と呼ぶのが通例です。 クリチン(繊維)が生理的静穏のステーションで休んでいる場合、内部電荷は音に対して負であり、精神的にゼロとして受け入れられます。 異なる組織では、膜電位が特徴づけられます 異なるサイズ:m'yazovoi組織で最も高い-80 -90 mV、神経組織で-70 mV、幸せな-35 -40 mV、上皮組織で-20mV。
Utvorennya MPPは、イオンK +、Na +、Ca 2+、Cl-の濃度、およびクリチンの芽の膜の特徴に存在します。 膜にあるイオンチャネルであるゾクレマは強力かもしれません:
1.選択性(選択的浸透)
2.電気。
穏やかな状態では、ナトリウムチャネルは閉じており、カリウムチャネルのほとんどは開いています。 チャネルは丸まって丸くなることがあります。 膜で チャネル(非特異的)、yakієはすべての要素に浸透しますが、カリウムにはより浸透します。 カリウムチャネルは常に振動しており、イオンは濃度と電気化学的勾配に沿ってqチャネルを介して崩壊しています。
膜イオン理論によれば、MPPの存在は次のように示されます。
Ø膜のイオンチャネル内のイオン循環によって中断されることなく、
Ø膜の両側で絶えず変化する陽イオンの濃度、
Øナトリウム-カリウムポンプの中断のない動作。
Øこれらのイオンのチャネルの浸透が異なります。
ІonіvK+はクリチナムが豊富で、ヨガの音は小さく、Na +-一方、位置はクリチナムが豊富で、クリチナムでは十分ではありません。 ІonіvСl–クリチンの名前よりも大きいトロッホ、中央下。 真ん中のセルは有機アニオンが豊富で、膜の内面の負電荷を保護するために重要です。
穏やかな細胞膜では、K+イオンのみが浸透します。 彼らは平和な状態でカリウムを持っています 途中で高濃度のNa+。 それに、落ち着くと膜の外面が正に帯電します。 高分子の有機アニオン(タンパク質)は膜の内面に集中しており、負の電荷を示します。 膜の外側からの悪臭は静電的にUtrimuyutK+イオンです。 MPPを照らす主な役割は、Do+イオンにあります。
チャネルを介してイオンが流れた後、イオンの濃度の差は変化しません。 不変の世話をします。 膜がNa+-最大+-ポンプを使用することは何もありません。 悪臭は細胞からNa+を継続的に放出し、反対の濃度勾配をK+細胞質に注入します。 細胞から採取された3つのNa+イオンの場合、2つのK+イオンが中央に導入されます。 濃度勾配に逆らうイオンの移動は、(大量のエネルギーからの)能動輸送によって引き起こされます。 ATPエネルギーが存在するときは、gyne。
落ち着きの可能性の現れは、弱者を分割した後、クライアントが機能的な落ち着きの状態から目覚めの状態に移行することを実質的に軽減することを可能にします。
励起されると、再充電による膜の静止電位の大きさが減少します。 膜の内部電荷が負にならなくなると、膜の脱分極が起こり、diの電位が発生し始めます。
4.DIYとメカニズムのヨガ旅行の可能性。
目覚めのSpivvіdshenniaフェーズと潜在的なdiїのフェーズ。
潜在的な神経、m'yazovyhおよび分泌クリチンの目覚めのせいであるshvidke膜電位に名前を付けます。 ヨガの基礎には、膜のイオン透過性の変化があります。 電位の変化の振幅と性質は、ティーザーの強さにあるのに十分ではありません。ティーザーの強さは、ブラの強さが臨界値以上になるように、より重要です。からかいのしきい値と呼ばれます。
中断しきい値--tsya最小の力、障害がある場合、力の反作用は最小です。 razdratuvannyavikoristovuyutのしきい値を特徴づけるために理解する レオベース(Reo-ストラム、ベース-メイン)。
スレッショルドクリームは分割されています サブスレッショルドpodrazniki、 yakіは声の反応を要求することができます、さもなければクリチンのスピーチの交換の破壊を要求することができます。 それで しきい値を超えるpodrazniki。
Vyniknuvshi、PDは膜を拡張し、 変わらない独自の振幅。 新しいフェーズが分離されます:
1)脱分極:
a)完全な脱分極;
b)shvidka脱分極。
2)再分極:
a)遅い再分極;
b)完全な再分極(負のトレース電位)
3)過分極(正の微量電位)
選択的に(1種類のイオンのみを透過します)。 イオンの性質上、悪臭はNa +、Ca ++、Cl-、K+チャネルを通過させます。
非選択的(多くの種類のイオンに浸透します);
2)規制の方法によると、それらは次のように分けられます。
潜在的な預金(電気的励起、増強)
潜在的な独立(化学活性、(リガンド-受容体-沈着)、ケモケロバニ)
メカノエキサイティング(機械工学)。
安心の可能性。 心の安らぎの可能性に関する膜イオナ理論。 Mіstseve私はより広いzbudzhennya。

生細胞の膜は分極しており、内面は正常に対して電気陰性であることが確立されています。 膜電位は良好です–(マイナス)70 –(90)mV。 励起されると、再充電による膜の静止電位の大きさが減少します。 この節約ポテンシャルの形成は、膜のイオンチャネルに沿ったイオンの中断のない循環、膜の両側からの陽イオンの濃度の一定の差、ナトリウム-カリウムポンプの中断のない動作によって穏やかに妨げられます。 イオン濃度の違いと膜の分極は、ナトリウムイオンが絶えず排出され、カリウムイオンが細胞に活発に移動する間維持されます。 細胞内のカリウムイオンの濃度は、クリチン後の濃度を30〜40倍上回り、ナトリウムのクリチン後の濃度は、クリチン内の濃度よりも約1桁高くなります。 Електронегативність внутрішньої поверхні мембрани обумовлена наявністю в клітині надлишку аніонів органічних сполук, абсолютна величина потенціалу спокою (мембранний потенціал, трансмембранний потенціал, рівноважний калієвий потенціал) обумовлена головним чином співвідношенням внутрішньо- та позаклітинної концентрацій іонів калію та задовільно описується рівнянням ネルンスト:
 (1)
(1)
vrakhovuєの現代の理論は同じです:
1)ナトリウム、塩素、カルシウムイオンの濃度の違い。
2)1時間あたりの皮膚イオンの膜の浸透(P)。
落ち着きの可能性の現れは、弱者を分割した後、クライアントが機能的な落ち着きの状態から目覚めの状態に移行することを実質的に軽減することを可能にします。
Viniknennyaの可能性dії(脱分極)

I-膜電位
1-静止電位、2-完全な脱分極、3-遅い脱分極、4-遅い再分極、5-完全な再分極、6-過分極
II-覚醒
a-通常、b-高度、-絶対的な不応性、
d –目に見える不応性、e –超正常性、
e-非正規性
電位dії(PD)は、膜の見かけの分極(電位の落ち着き)により発生し、イオンチャネル(ナトリウムおよびカリウム)の透過性を変化させます。 電位が変化するとすぐに、チャネルの活性化によりイオンの浸透が増加します。 ナトリウム、これはクライアントに入り、脱分極のプロセスを確実にします。 ナトリウムイオンのクリチンへの入場は、膜の内面の電気陰性度を変化させ、それにより、新しいイオン性ナトリウムチャネルの活性化およびクリチンへのナトリウムイオンのさらなる流入が可能になる。 力:
a)内部の細胞陰イオン基の静電重力;
b)ナトリウムイオンの濃度勾配、セルの中央をまっすぐにします。
クリチン内のナトリウムイオンの等電荷と、イオンの帯電と同時に空気の力の流入下での等距離の目がくらむ可能性のピーク。
生体膜は、細胞質と内部クリチン構造の大部分を取り囲む機能的に活性なクリチンの構造です。 団結を確立する 内部クリチンシステム細管、折り目、閉じた空きスペース。
膜の構造的基盤は、膜タンパク質を含むリン脂質の下部球です。 クリチン膜の厚さは6〜12nmです。 脂質分子は両性です。 その親水性の部分で、獣の悪臭は水媒体(細胞間組織と細胞質)の近くにあり、分子の疎水性の部分はリン脂質ビスルの中央に向けられています。 この構造は、クリチン後相とクリチン内相の分割に最適です。
極性細胞によってリン脂質のアンダーボールに注入されたタンパク質は、水相の近くに親水性の表面を確立します。 悪臭 さまざまな機能:受容体、酵素、イオンチャネル、膜ポンプ、およびイオンと分子の担体。
イオンチャネルのパワーに関する私たちの知識のほとんどは、機能の再構築の結果です。 スキンチャネルmaє女の子、選択フィルター、ゲートおよびkeruvannyaゲートのメカニズム。
チャネルの一部は電気的にエッチングされています。 膜上の電位の分布を管理します(電位沈着イオンチャネル)。 チャネルの最初の注文には、膜電位の値に依存する電気センサーがあります。これは、チャネルのゲートを開くか、閉じるかのいずれかです。
イオンチャネルの別のオプションは、受容体の角質化です。 ゲートは、膜の表面に広がる受容体遺伝子座によって制御されます(メディエーターが受容体と相互作用すると、受容体は開きます)。 受容体と一般的なメカニズムの間のいくつかの受容体-セラミックチャネルには、中間段階があります(中間cAMPタイプ、プロテインキナーゼなど)。
これらのチャネルは、選択性と導電性という2つの重要な膜力を提供します。
チャネルの選択性または活気は、その特別なタンパク質構造、チャネルの形状によって保証されます。
たとえば、ナトリウムイオンの直径は0.19 nmで、ワインの水和シェルと合わせて0.3nmに近くなります。 ナトリウムチャネルのネック0.3〜0.5nm。 チャネルを通過するために(特に選択フィルターを通過するために)、ナトリウムイオンまたは別のイオンは水和膜の形であるという罪を犯し、「裸の」見た目の人がチャネルを通過する可能性は低くなります。 大きすぎるイオンは首に入ることができず、小さすぎる、不自然なイオンはハイドレーションシェルを首に入れることができません 選択フィルターチャンネルから抜け出すことはできません。
ナトリウムチャネル(図6)は、活性化(mゲート)と不活性化(hゲート)の2種類のゲートを操作します。 穏やかな活性化ゲートの心の中で閉じられていますが、軽減する準備ができており、不活性化ゲートは準備ができています。 MPが低下すると(60 mVへの脱分極)、活性化ゲートが開き、ナトリウムイオンがクリチナに流入し、次に不活性化ゲートが閉じ始めます(ナトリウムチャネルの不活性化)。 1時間後、アクティブ化ゲートが開始され、非アクティブ化ゲートが開き、チャネルは新しいサイクルの準備が整います。 チャネルは、テトロドトキシン、局所麻酔薬(ノボカインおよびイン)によって遮断されます。
米。 6.ナトリウムチャネルと「ゲートウェイ」メカニズムの働き。
A-落ち着いて、mゲートは閉じられます。 B-zbudzhennym-gatevіdkritіの場合; B-脱分極中のh-voritの閉鎖(不活性化)。
カリウムチャネルは選択的に投与することもできます-カリウムイオンを通過させることが重要です。 テトラエチルモニウムによってブロックされています。 悪臭の不活化のプロセスは弱く表現されています。 次に、細胞内カルシウム濃度が上昇し、細胞膜の脱分極が起こると活性化される特別なカリウムチャネルがあります。 カリウム-カルシウム沈着チャネルの活性化は、再分極を加速します(回復性MP)。
カルシウムチャネル。 入口カルシウムストラムは、通常クリチン膜を脱分極させるのに十分な大きさではありません。 カルシウムは二次として機能します
仲介者(メッセンジャー)。 カルシウムチャネルの活性化は、例えばナトリウム入口流による細胞膜の脱分極によって確実になります。 カルシウムチャネルの不活性化は、遊離カルシウムの内部細胞濃度の増加によるものです。 細胞質のタンパク質タンパク質はカルシウムに結合します。これにより、1時間で、低レベルでも安定した量のカルシウムストローマを維持できます。 このナトリウムストラムを使用すると、より敬意を払うようになります。 カルシウムチャネルは、マンガン、ニッケル、カドミウムのイオン(2価イオン)、および薬用スピーチ(ベラパミル)によって遮断されます。
膜を通過するイオンの受動的(エネルギーフリー)輸送と能動的(エネルギー貯蔵)輸送を区別します。
単純で容易な拡散の説明のために受動的に行きます。
単純拡散は、化学的、電気化学的、または浸透圧勾配の背後にあるフィックの法則に従います。 たとえば、クリチンには14ミリモルのナトリウムが含まれ、中央には140ミリモルのナトリウムが含まれています。
受動的な単純拡散の場合、発話は脂肪になる可能性があります。 親水性のスピーチは、膜のリン脂質バイスフィアを通過できません。
拡散は、輸送される分子に特異的に結合し、濃度勾配に沿った移動に付着するため、特別なチャネルの存在またはキャリアの関与のいずれかによって促進されます。
クリチン膜のポンプによるイオンの能動輸送は、膜の両側からのイオン勾配のサポートを保証します。 エネルギーは、その集中の勾配に対するスピーチの伝達に反映されます。
特殊な酵素システムでのイオンの能動輸送への参加-ATPアーゼ、ATPのyakіzdіysnyuyut加水分解。 別:
ナトリウム-カリウム-ATP-アーゼ(「ナトリウムポンプ」)は、すべての生き物、ロスリン、微生物のクリチンの膜に含まれています。 これは、イオンの2つの結合中心を持つことができる膜タンパク質です。 それらの1つ(ナトリウム)はクリチン膜の内面に広がり、もう1つ(カリウム)は外面に広がります。 強心配糖体酵素の特異的阻害剤は、ナトリウムポンプを遮断するストロファンチン(ウアバイン)です。 1つのATP分子の加水分解には、細胞からの3つのナトリウムイオンの除去と、細胞による2つのカリウムイオンの注入が伴います。 ポンプの中央セルのカリウムイオンまたは中央セルのナトリウムイオンの量が増えると、ポンプがアシストされます。
カルシウム-ATP-アーゼ(「カルシウムポンプ」)は、悪性細胞の筋小胞体の膜で最も広く見られます。
プロトン-ATP-アーゼ(「プロトンポンプ」)-ミトコンドリアの膜にあります。